Embryo series courtesy of Einhard Schierenberg
Embryo series courtesy of Einhard SchierenbergTable of Contents
Abstract
Cell-cell interactions mediated by the Notch signaling pathway occur throughout C. elegans embryogenesis. These interactions have major roles in specifying cell fates and in tissue morphogenesis. The network of Notch interactions is linked in part through the Notch-regulated expression of components of the pathway, allowing one interaction to pattern subsequent ones. The Notch signal transduction pathway is highly conserved in animal embryogenesis. The REF-1 family of bHLH transcription factors are major targets of Notch signaling in the C. elegans embryo, and are distantly related to HES proteins that are targets of Notch signaling in Drosophila and vertebrates.
The Notch signaling pathway plays a central role in patterning metazoan development, and is used in remarkably diverse cell fate decisions (for general review, see Artavanis-Tsakonas et al., 1999). C. elegans contains two, closely-related proteins called GLP-1 and LIN-12 in the Notch family of transmembrane receptors. Cell interactions mediated by these receptors have been documented throughout embryonic and postembryonic development of C. elegans. For example, Notch signaling controls mesoderm induction during embryonic development, and controls germ cell mitosis during postembryonic development (see below and Austin and Kimble, 1987).
The general mechanism of signal transduction, here called the Notch pathway for simplicity, has been elucidated primarily through studies in C. elegans and Drosophila, and is reviewed in detail in LIN-12/Notch signaling in C. elegans. Briefly, signaling is activated when the receptor Notch contacts a ligand in the DSL (Delta/Serrate/LAG-2) protein family. Inductive interactions occur when one cell expresses the receptor and a neighboring cell expresses the ligand. In lateral signaling, two or more equivalent cells express both the ligand and the receptor; interactions between the cells modulate ligand/receptor levels to achieve asymmetry (see Wilkinson et al., 1994). Contact between ligand and receptor initiates a series of cleavage events that liberate the intracellular domain of the receptor. This domain then enters the nucleus, where it activates gene expression in conjunction with the DNA-binding protein LAG-1/Su(H).
A principal target of Notch signaling in the embryo is the ref-1 gene family, consisting of ref-1, hlh-25, hlh-26, hlh-27, hlh-28, and hlh-29 (Neves and Priess, 2005). One or more of these genes is expressed after each of the Notch interactions in the early embryo. These genes encode unusual proteins with two separate bHLH (basic-helix-loop-helix) domains (Alper and Kenyon, 2001). Each basic domain shows moderate similarity to the basic domains of HES (Hairy and Enhancer of Split) proteins that are principal targets of Notch signaling in Drosophila and vertebrates. HES proteins mediate transcriptional repression by binding the corepressor Groucho; REF-1 has been shown to bind C. elegans UNC-37/Groucho, and depleting UNC-37 can cause defects resembling those caused by depleting REF-1 family members (Neves and Priess, 2005). However, the REF-1 proteins lack several key characteristics of HES proteins, and thus appear to be highly diverged relatives. The network of Notch interactions in the C. elegans embryo suggests that there are several additional, as yet unidentified, targets of Notch signaling. Recent studies have identified a group of Notch targets during postembryonic development, but it is not known whether any of these function in the embryonic interactions (Berset et al., 2001; Gupta and Sternberg, 2002; Lamont et al., 2004; Yoo et al., 2004).
The first division of the fertilized egg produces two cells called AB and P1 (Figure 1). These cells undergo very different, but reproducible, patterns of division and differentiation that are referred to as cell lineages (Sulston et al., 1983). The division of P1 is oriented along the anterior/posterior axis; this division is asymmetric and produces daughters (EMS and P2) that express distinct sets of proteins. For example, only P2 expresses the Notch ligand APX-1/Delta (Mickey et al., 1996). The division of AB is oriented along the transverse axis, but as the spindle elongates one AB daughter is displaced toward the posterior; this daughter is called ABp and the more anterior daughter is called ABa. ABa and ABp initially are equivalent (Priess and Thomson, 1987), and both express the receptor GLP-1/Notch (Evans et al., 1994) (see also Translational control of maternal RNAs). However, the posterior displacement of ABp puts it in contact with the ligand-expressing P2 cell; thus in a 4-cell embryo GLP-1/Notch is activated in ABp, but not in ABa (Figure 1). The Notch signal transduction components that mediate the first interaction, such as GLP-1 and APX-1, are expressed maternally (Mango et al., 1994; Mello et al., 1994). Notch signaling induces the embryonic expression of the ref-1 family within about 25 minutes, corresponding to the birth of the ABp granddaughters (Figure 2; Neves and Priess, 2005).
|
Figure 1. Schematic diagram of early blastomeres at the 2-cell, 4-cell, and 12-cell stages; the 12-cell embryo is a ventral view with the AB descendants splayed to demonstrate cell contacts. For simplicity, AB descendants at the 12-cell stage such as ABala or ABarp are labeled "ala" and "arp". In this and other Figures, cells expressing GLP-1/Notch are outlined in red, and Notch-activated cells are shown first in light green, then dark green at later time points.
At the 12-cell stage of embryogenesis, maternally-expressed GLP-1/Notch remains on the surfaces of the ABa descendants, and two new P1 descendants, called MS and E, become signaling cells (Figure 1; Hutter and Schnabel, 1994; Mango et al., 1994; Lin et al., 1995). Genetic studies suggest that the signal(s) expressed by MS and E are the products of embryonically-transcribed genes, because signaling is dependent on the transcription factor SKN-1 (Bowerman et al., 1992; Shelton and Bowerman, 1996). Although the molecular identify of the MS signal is not known, it is likely to be a Delta-related ligand similar to APX-1 because P2 can partially substitute for MS signaling within chimeric embryos (Shelton and Bowerman,1996). MS contacts two of the four ABa descendants present at the 12-cell stage, and activates GLP-1. The entire ref-1 family is expressed about 25 minutes later in the granddaughters of those two ABa descendants (Figure 2, see also Figure 4; Neves and Priess, 2005). ABp descendants also continue to express GLP-1/Notch at the 12-cell stage, and these cells also are in contact with MS or E. However, the Notch-activated ABp granddaughters appear to be refractive to the second Notch interaction in normal development, and can respond only if the first Notch interaction is blocked by manipulation or mutation (Mello et al.,1994; Moskowitz et al., 1994).
|
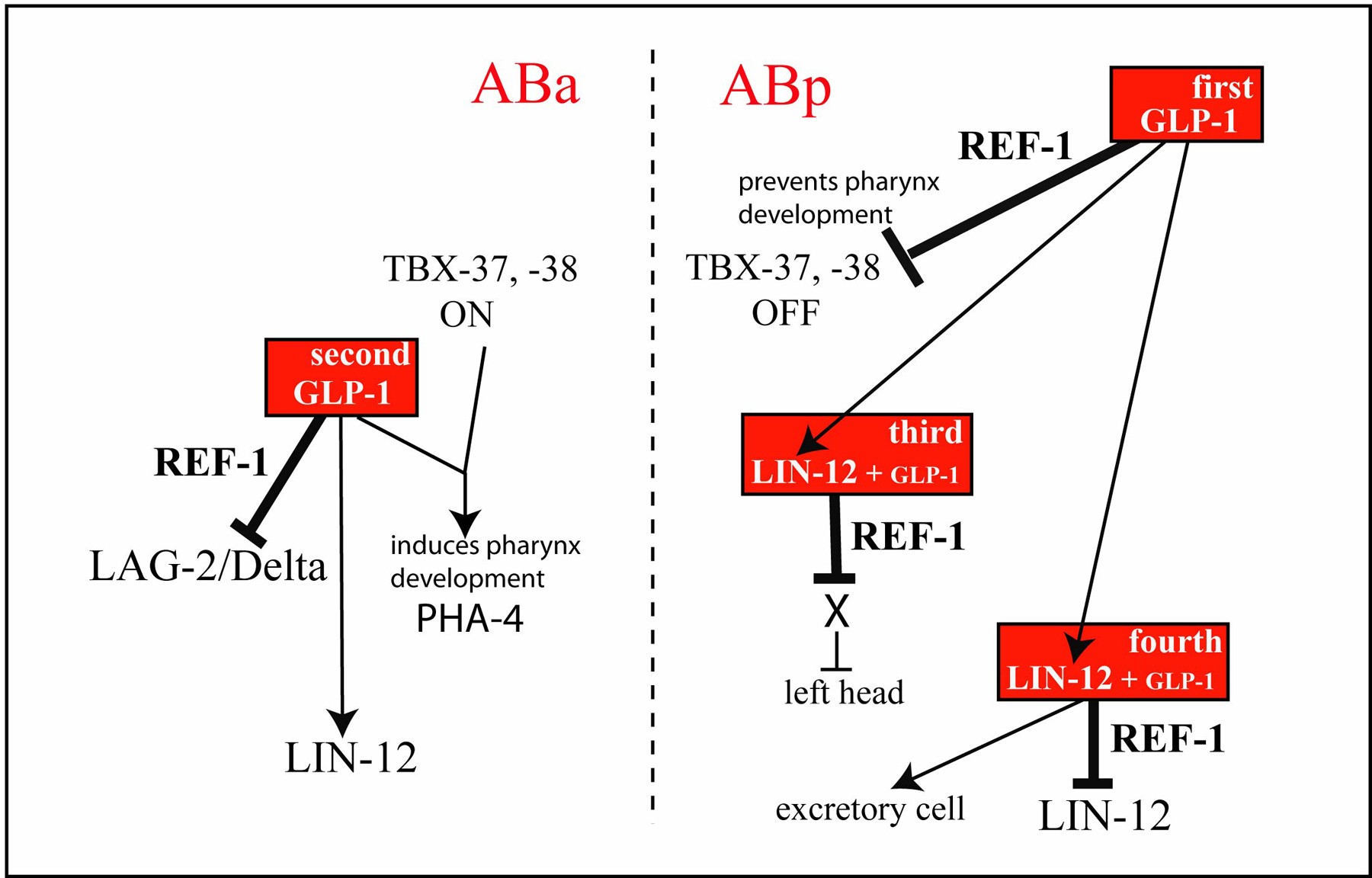
Figure 2. The first four Notch interactions in the AB lineage. A partial lineage of the AB blastomere is shown in red; for simplicity the vertical axis is not scaled accurately with respect to time. A subset of descendants of ABa and ABp that express GLP-1 or LIN-12 contact various ligand-expressing cells (blue) and activate Notch signaling. Expression of the REF-1 family members occurs about 25 minutes after an interaction (bold black lines). For comparison with Figure 5, the left and right head precursors are indicated. The bottom panel shows the effects of Notch signaling on the various fates of AB descendants. The pattern of cell division and differentiation of a cell is considered here as its 'fate'. In wild-type development, the eight AB descendants have unique fates that can be represented by their names (ABala, ABarp, etc); for simplicity, these different fates/names are represented as numbers with the key shown at right. Defects in Notch signaling transform one or more of these cells to resemble other cells as shown. In the absence of Notch signaling, all eight cells express TBX-37, -38 (magenta) and adopt one of two fates (1a or 1p, where a= anterior and p=posterior). For example, all of the 1a cells adopt a pattern of development that resembles a wild-type ABala cell. When only the first interaction occurs, TBX-37, -38 are repressed in ABp descendants and these cells adopt new fates as shown. If embryos undergo both the first and second interaction, ABa descendants (boxed) also change their fate as indicated. When all four interactions occur (bottom line), all AB descendants have their wild-type fates.
One role of the first and second AB interactions is to define which AB descendants produce pharyngeal cells (Figure 3). The C. elegans pharynx is primarily a mesodermal organ containing muscle cells, gland cells, support cells, and several neurons. The key regulator of pharyngeal development is the forkhead transcription factor PHA-4 (Mango et al., 1994; Azzaria et al., 1996; Horner et al., 1998; Kalb et al., 1998). PHA-4 is essential for the development of all pharyngeal cell types (Mango et al., 1994), and forced expression of PHA-4 is sufficient to induce ectopic pharyngeal development (Horner et al., 1998; Kalb et al., 1998). In early embryogenesis, the Notch-activated ABa descendants, but not ABp descendants, express PHA-4 and produce pharyngeal cells. Preventing the second Notch interaction by depleting Notch pathway components such as GLP-1, APH-1, APH-2, or SEL-8 (Priess et al., 1987; Doyle et al., 2000; Goutte et al., 2000; Petcherski and Kimble, 2000; Goutte et al., 2002), or by killing the signaling cell MS (Hutter and Schnabel, 1994; Mango et al., 1994), prevents ABa descendants from producing pharyngeal cells. Conversely, preventing the first interaction allows ABp descendants to respond to signaling from MS (and E), such that these descendants produce pharyngeal cells ectopically (Hutter and Schnabel, 1994; Mello et al., 1994; Moskowitz et al., 1994). Thus in normal development the first interaction prevents, and the second interaction induces, pharyngeal development.
|
Figure 3. Model summarizing the various outputs of the first four Notch interactions in the AB lineage. The first two interactions are mediated by GLP-1, and the second two require either LIN-12 and GLP-1. For example, the second interaction (mediated by GLP-1) activates the expression of LIN-12/Notch, but represses the expression of LAG-2/Delta. Although the tbx-37, -38 genes appear to be direct targets of REF-1 family members, it is not known whether other examples of repression are direct or indirect. Adapted from Neves and Priess (2005).
Because the lack of induced pharyngeal tissue, (caused by a defect in the second interaction), can mask the hyperinduction of pharyngeal tissue (caused by a defect in the first interaction), the role of GLP-1 in the first interaction was discovered only after the second interaction had been well-characterized. Early experiments demonstrated that interactions between ABp and P2 at the 4-cell stage were required for ABp to produce a non-pharyngeal tissue (valve cells; Bowerman et al., 1992). However, the requirement for GLP-1 in the interaction between ABp and P2 was deduced from later discoveries that (1) the P2 ligand was a Delta-like protein, (2) temperature-sensitive GLP-1 alleles had an additional temperature-sensitive period before the 12-cell stage, (3) glp-1 mutants had lineage defects in ABp as well as in ABa descendants, and that (4) preventing P2 from contacting ABp in wild-type embryos caused lineage defects similar to those seen in glp-1 mutants (Hutter and Schnabel, 1994; Mello et al., 1994; Moskowitz et al. 1994).
Why do two Notch-mediated interactions that occur in rapid succession at the 4-cell and 12-cell stages, that use the same receptor (GLP-1) and functionally similar ligands, and that have at least one common target (the ref-1 family), have opposite effects on pharyngeal development? Cell culture experiments suggested that the age of the AB descendants is critical for determining the respective outcomes of the two interactions (Shelton and Bowerman, 1996). In these experiments, an AB blastomere was isolated and allowed to divide in culture to the equivalent of the 12-cell stage, then combined with a P2 blastomere from a 4-cell embryo. The AB descendants responded by expressing pharyngeal markers appropriate for the 12-cell interaction, rather than markers appropriate for the 4-cell interaction. The molecular basis for this time-dependent difference appears to be two functionally-redundant T-box transcription factors called TBX-37 and TBX-38 that are essential for AB descendants to produce pharyngeal tissue (Good et al., 2004). The first Notch interaction represses tbx-37, -38 expression in ABp descendants, thus preventing those cells from producing pharyngeal tissue (Figure 2, Figure 3 and Figure 4). Indeed, repression of tbx-37, -38 appears to be a primary function of the first Notch interaction: Many of the defects in ABp development that result from blocking the first Notch interaction can be suppressed by simultaneously removing TBX-37, -38 activities (Good et al., 2004). Thus the first interaction restricts the competence to produce pharyngeal cells to ABa descendants, by repressing the tbx-37, -38 genes, while the second Notch interaction specifies which of the ABa descendants that express TBX-37, -38 will produce pharyngeal tissue.
|
Figure 4. Ventral view of 26-cell embryos showing expression of TBX-38 and a REF-1 family member, HLH-29. At this stage there are eight ABa, and eight ABp, descendants. ABp descendants are induced to express HLH-29 and other REF-1 family members by the first Notch interaction, thus preventing TBX-38 expression in these cells. HLH-29 and a subset of other REF-1 family members are expressed in EMS descendants (circled) independent of Notch, and similarly prevent TBX-38 expression. TBX-38 is expressed at high levels in all ABa descendants before the second Notch interaction induces high levels of the REF-1 family. The asterisk indicates an example of an ABa descendant that expresses TBX-38 but is not activated by Notch signaling. Adapted from Neves and Priess (2005).
tbx-37, -38 appear to be direct targets of REF-1-mediated repression after the first Notch interaction. Depletion of REF-1 family members causes derepression in ABp descendants of a transgene containing sequences upstream of tbx-37, and REF-1 can bind an element from the tbx-37 promoter in vitro (Neves and Priess, 2005). Although the ref-1 family is expressed in response to Notch activation in the AB lineage, some members of the ref-1 family are expressed in EMS descendants (Robertson et al., 2004; Broitman-Maduro et al., 2005) in a pathway that is independent of Notch signaling (Figure 4; Neves and Priess, 2005). This Notch-independent pathway involves the maternally-expressed transcription factor SKN-1 (Neves and Priess, 2005) and embryonically-expressed targets of SKN-1 called MED-1, -2 (Maduro et al., 2001; Broitman-Maduro et al., 2005). MED-1,2 are related to GATA transcription factors, but recognize non-canonical sites in the promoters of some ref-1 family members (Broitman-Maduro et al., 2005). Depletion of REF-1 family members derepresses tbx-37, -38 expression in EMS descendents, suggesting the REF-1 family has similar functions in ABp and EMS cells irrespective of how ref-1 expression is initiated (Neves and Priess, 2005).
If the REF-1 family members repress tbx-37, -38 in ABp and EMS descendants, why do REF-1 family members fail to repress tbx-37, -38 in the Notch-activated ABa descendants? The timing of the Notch interactions relative to tbx-37, -38 expression may be the critical difference between these cells. At the 24-cell stage, when TBX-37, -38 is first detectable, the REF-1 family is present at high levels in ABp and EMS descendants, but present at only low levels in the Notch-activated ABa descendants (Figure 4; Neves and Priess, 2005). Thus there may be insufficient levels of the REF-1 family members in ABa descendants to prevent TBX-37, -38 expression. The target(s) of the second Notch interaction that collaborates with TBX-37, -38 to induce expression of PHA-4, and thus promote mesodermal development, has not yet been identified.
In normal development, an ABp descendant called ABplaaa and an ABa descendant called ABarpap produce cells that contribute to the left and right sides of the head, respectively (Sulston et al., 1983). The divisions and differentiation patterns of the left and right head precursors are identical, although the sisters and cousins of these cells have very different developmental patterns. The first evidence that cell interactions were involved in head development came from the finding that lin-12 glp-1 double mutants had a 'twisted nose' (Lambie and Kimble, 1991). The developmental pattern of the right head precursor is not noticeably altered by killing neighboring cells, suggesting that it is not influenced by cell interactions . However the development of the left head precursor is altered markedly when precursors of neighboring cells are killed (Hutter and Schnabel, 1995a; Moskowitz and Rothman, 1996). Mutations in genes encoding the Notch pathway components LAG-2/Delta, LAG-1/S(uH), or LIN-12/Notch together with GLP-1/Notch cause similar defects in ABplaaa development, indicating that the fate of the left precursor is specified by the Notch pathway (Figure 5; Moskowitz and Rothman, 1996). The left precursor expresses LIN-12, and appears to be signaled by a neighboring cell called ABalapp that expresses the ligand LAG-2 (Moskowitz and Rothman, 1996).
|
Figure 5. Notch signaling and head development. The top four panels show embryos as listed that were stained to visualize adherens junctions surrounding the hypodermal (skin) cells on the surface (see Epidermal morphogenesis). The right side of the lin-12 glp-1 mutant is similar to wild-type embryos; the H0 and H1 hypodermal cells produced by the right head precursor are indicated with asterisks. Note that these cells are not produced by the left head precursor in this embryo, nor are they produced on the left side of lag-1 or lag-2 mutant embryos. The bottom panels show a dorsal view of a wild-type embryo with the granddaughters of the left and right head precursors indicated. Both sets of granddaughters normally express REF-1, however killing the ligand-expressing cell for the third Notch interaction blocks expression only in the left cells. Adapted from Moskowitz and Rothman (1996); Neves and Priess (2005).
The ref-1 gene is a target of the third Notch interaction, and is expressed in the daughters and granddaughters of the left head precursor (Neves and Priess, 2005). Because the role of the third interaction is to make the left precursor identical to the right, it is intriguing that ref-1 is expressed simultaneously in the daughters and then granddaughters of the right head precursor; this expression pattern is independent of Notch (Figure 5). ref-1 mutants show defects on both the left and right sides of the head, suggesting that ref-1 must function on both sides for proper development (Alper and Kenyon, 2001; Neves and Priess, 2005). Thus the bilateral symmetry of the C. elegans head results in part from Notch-dependent REF-1 expression on the left side, and Notch-independent REF-1 expression on the right side.
The fourth Notch interaction involves an AB descendant called ABplpapp. Cell killing experiments and analysis of Notch pathway mutants showed that the fate of ABplpapp is specified through Notch signaling (Hutter and Schnabel, 1995a; Moskowitz and Rothman, 1996). In normal development one of the ABplpapp descendants produces the excretory cell (Sulston et al., 1983), a cell type missing in glp-1 lin-12 double mutants and in lag-2 or lag-1 single mutants (Lambie and Kimble, 1991). Cell killing experiments, and localization studies suggest that the Notch pathway is activated in ABplpapp through LAG-2 signaling from one or both of the daughters of a cell called MSap (Hutter and Schnabel, 1995a; Moskowitz and Rothman, 1996). ref-1 is a target of the fourth interaction, and appears in the daughters of ABplpapp (Figure 2; Neves and Priess, 2005). Because other ref-1 family members do not appear to be expressed in these cells, and ref-1 mutants have an excretory cell, there may be additional Notch targets in the fourth interaction.
AB descendants normally have unique developmental patterns that can be called 'fates'. The ability to trace division and differentiation patterns (lineages) of individual cells in living C. elegans embryos provides a wealth of information for assigning cell fate. A typical cell lineage might contain asymmetric divisions, programmed cell deaths, and descendants that undergo characteristic numbers of cell cycles before terminally differentiating. For example, cells like ABarp and ABprp are both ectodermal precursors that produce some descendants that express similar molecular markers. However, the lineages of these cells are very different, and it is much more informative to say that ABarp has an 'ABarp fate' than to say it is an ectodermal precursor.
Although Notch interactions occur both before and after the AB8 stage, when there are eight AB descendants. This stage provides a useful reference point for summarizing the effects of Notch signaling on the fates of AB descendants (Figure 2). After the first four Notch interactions are completed in wild-type embryos, there are only two AB8 cells that have not experienced Notch signaling; these are the cells ABala and ABarp (Figure 1 and Figure 2). As cell lineages in mutants defective in the Notch pathway were determined, it was discovered that multiple AB8 descendants were transformed such that they acquired characteristics of wild-type ABala and ABarp cells (Hutter and Schnabel, 1994; Mello et al., 1994; Moskowitz et al., 1994). The most extensive of these lineage studies (Hutter and Schnabel, 1994) showed that the transformations were remarkably complete. Thus, the ABala and ABarp lineage patterns can be considered as primary fates for all AB8 cells, with the four Notch interactions diversifying these fates.
As shown in the summary model in Figure 2 (see also Hutter and Schnabel, 1994; Moskowitz et al., 1994; Hutter and Schnabel, 1995a), the AB8 cells consist of two pairs of anterior/posterior sister cells from ABa and two pairs from ABp. In the absence of Notch signaling, each of the four anterior sister cells adopts a primary fate (represented by 1a= ABala) and each of the four posterior sister cells adopts a different primary fate (represented by 1p = ABarp). TBX-37, -38 contribute to these primary fates in normal development, and a failure in the first interaction causes misexpression of TBX-37, -38 in ABp descendants (Good et al., 2004). In the descendants of ABp, the first Notch interaction converts primary fates 1a and 1p into secondary fates 2a and 2p, respectively, by repressing expression of TBX-37, -38. The second Notch interaction, in ABa descendants, converts 1a and 1p fates into 3a and 3p fates, respectively; in these cells TBX-37, -38 collaborate with an unknown target of Notch signaling to induce pharyngeal development (Good et al., 2004). The third and fourth interactions convert 2a into 4a fates, and 2p into 5p fates, respectively. This simple code for the first four Notch interactions predicts most of the cell lineage changes observed in embryos that either lack specific Notch interactions, or that undergo ectopic Notch interactions (Hutter and Schnabel, 1994; Moskowitz et al., 1994; Hutter and Schnabel, 1995a).
The model for the first four Notch interactions requires that there is a Notch-independent difference between anterior and posterior sister cells (1a vs 1p, 2a vs 2p, etc). Several early studies provided evidence for differences between anterior and posterior sister cells in C. elegans, and suggested that cells throughout the embryo somehow recognized a common anterior/posterior axis of polarity (Way et al., 1994). For example, several anterior/posterior divisions occur in late embryogenesis that show similar, asymmetrical expression of the transcription factor UNC-86 (Finney and Ruvkun, 1990) Analysis of mutants with transformed cell lineages in early embryogenesis suggested that early cells also recognize a common axis of anterior/posterior polarity, irrespective of cell fate (Mello et al., 1992). For example, the MS blastomere is positioned in the middle of a wild-type embryo, and the MS descendant born from the cleavage pattern posterior/anterior/anterior/posterior/posterior (MSpaapp) invariably undergoes the first apoptotic cell death; no other MS descendants die at that stage (Sulston et al., 1983). The developmental pattern of MS is determined in part by the transcription factor SKN-1 (Bowerman et al., 1992), and mutants with inappropriate SKN-1 activity in AB or P1 descendants produce ectopic-MS-like cells in the anterior and posterior of the embryo, respectively (Mello et al., 1992). Remarkably, the anterior and posterior MS-like cells can each produce a descendant from a paapp division sequence that undergoes apoptosis, as does MSpaapp in the middle of wild-type embryos. Thus, cells that are located in different positions, but that express the same transcription factor, sense anterior/posterior polarity in a similar manner and differentiate accordingly.
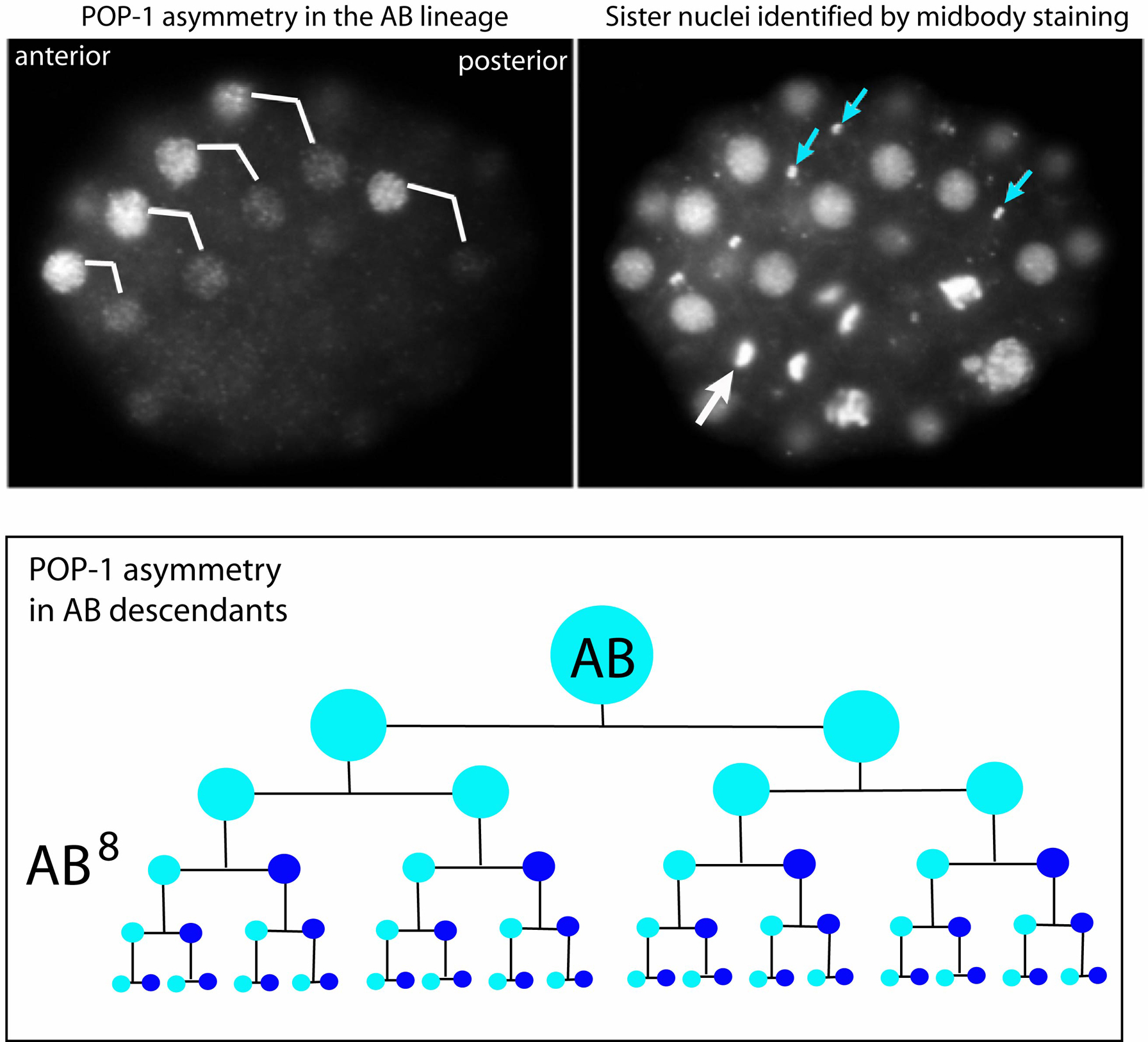
The transcription factor POP-1 appears to be the basis for the shared anterior/posterior polarity of embryonic cells; POP-1 is related to vertebrate Tcf/Lef proteins, whose activities are regulated by Wnt signaling (Lin et al., 1995; Kaletta et al., 1997; Lin et al., 1998). When an embryonic cell divides in C. elegans, the anterior daughter has a high level of POP-1 while the posterior daughter has a low level (Figure 6). In the AB lineage, POP-1 asymmetry first appears in the AB8 descendants, and is reiterated in successive divisions of these cells (Figure 6, lower panel). In numerous examples of anterior/posterior cell fate decisions, including the 1a/1p [ABala vs ABarp] decision, high levels of POP-1 are required for anterior fates (Lin et al., 1998). This suggests that POP-1 levels must be lowered for the posterior fate. POP-1 asymmetry is generated in the early embryo by a non-canonical Wnt pathway involving the ligand MOM-2/Wnt and a Nemo-like kinase called LIT-1 (Kaletta et al., 1997; Rocheleau et al., 1997; Thorpe et al., 1997; Rocheleau et al., 1999). The difference between anterior and posterior sisters appears to be generated by controlling the nuclear uptake of POP-1 after cell division (Rocheleau et al., 1999; Maduro et al., 2002; Lo et al., 2004).
Some experiments have suggested that AB descendants have an intrinsic ability to produce ABala-like cells (anterior fates), but that signaling between AB and P1 at the 2-cell stage initiates a latent polarity required for subsequent AB descendants to produce ABarp-like cells (posterior fates; Hutter and Schnabel, 1995b). However, other studies found that AB descendants showed characteristics of both ABala and ABarp development in the absence of P1 (Gendreau et al., 1994; Wittmann et al., 1997; Park and Priess, 2003). Although these differing results have not been entirely resolved, recent studies have suggested that POP-1 asymmetry at the AB8 and later stages may be generated by different pathways (Park and Priess, 2003). The normal onset of POP-1 asymmetry (at the AB8 stage) appears to involve MOM-2/Wnt signaling from P1 descendants; AB8 cells produced from an isolated AB blastomere do not show POP-1 asymmetry, while AB8 cells combined with P1 descendants exhibit POP-1 asymmetry. However, there appears to be a transition in later embryogenesis such that older cells can divide with POP-1 asymmetry in the absence of signaling from P1 descendants. This latter asymmetry does not require MOM-2/Wnt, but requires the transmembrane receptor MOM-5/Frizzled, a putative receptor for Wnt signaling. Interestingly, MOM-5::GFP shows an asymmetrical localization to the posterior pole of dividing AB descendants in normal embryos, and localizes asymmetrically in cultured, isolated cells that have no apparent Wnt signaling (Park et al., 2004). The correlation between asymmetric, high levels of MOM-5::GFP and the subsequent low level of nuclear POP-1 suggests that MOM-5/Frizzled asymmetry plays a role in normal POP-1 asymmetry. Although POP-1 asymmetry is evident after the AB8 stage in mutants lacking MOM-2/Wnt, cells do not show the normal anterior/posterior polarity of POP-1 expression. Thus early MOM-2/Wnt signaling may serve in part to orient the polarity of subsequent asymmetric cell divisions. The asymmetric localization of MOM-5::GFP is reminiscent of the Frizzled protein in Drosophila, which may be asymmetrically localized through the planar polarity pathway (Strutt, 2001). Future studies should address whether a similar pathway functions in the early C. elegans embryo.
|
Figure 6. POP-1 asymmetry in AB descendant. The left panel shows POP-1 expression in an embryo containing 32 AB descendants, and the right panel shows the same embryo stained with DAPI and an antibody that recognizes the midbody between sister cells (blue arrows). Note the high level of nuclear POP-1 in the most anterior sister of each pair in the left panel. The nuclear localization of POP-1 is cell cycle dependent and not seen during mitosis (large arrow in right panel indicates a dividing cell). The bottom panel shows a lineage diagram of AB descendants, with high nuclear POP-1 indicated by light blue and low nuclear POP-1 indicated by dark blue. Only the first few divisions of the AB descendants are shown. Adapted from Park and Priess (2003).
lin-12 mutants and lin-12 glp-1 double mutant embryos have several defects in AB development beyond those described above, indicating that there are multiple additional roles for Notch signaling in embryogenesis. The double mutants do not form a rectum and lack rectal cells (called K, K' and F, U), they lack the anal depressor muscle, and lack at least one intestinal muscle (Lambie and Kimble, 1991). The transcription factor PAL-1 appears to be required for rectal development (Edgar et al., 2001) and appears to be a direct target of Notch signaling (L. Edgar and B. Wood, unpublished). In wild-type embryos, PAL-1 is expressed in ABplpappp, the grandparent of the K and K' cells, and in ABplppppp, the grandparent of the anal depressor muscle and an intestinal muscle (Edgar et al., 2001). ABplpappp is a daughter of ABplpapp, the cell signaled by the fourth Notch interaction, however ABplppppp is not a descendant of ABplpapp. Thus PAL-1 expression in ABplppppp appears to be regulated by a fifth Notch interaction. Interestingly, the signaling cells for this interaction appear to be descendants of MSapa and MSapp (Edgar and Wood, unpublished), the same cells that function as signaling cells in the fourth AB interaction and the E4 interaction in the intestine (see below and Figure 7). The MSapa and MSapp cells express the ligand LAG-2/Delta while their bilateral symmetrical relatives do not (Moskowitz and Rothman, 1996). They enter the body cavity during gastrulation, where they contact the intestinal precursors and a succession of AB descendants that move toward the ventral midline during gastrulation (Figure 7 and see Gastrulation in C. elegans). Thus the MS descendants appear to be one of the main signaling centers in the embryo.
|
Figure 7. MSap descendants provide an inductive focus of Notch signaling on the left side of the embryo. Schematic diagram of a cross section through an embryo at successive time points. MS descendants are indicated with a bold black outline with the left, ligand-expressing cells shown in blue. LIN-12-expressing cells are outlined in red, and cells that undergo Notch signaling are filled with green. As gastrulation movements cause AB descendants to flow toward the ventral midline (arrows outside embryo), they come into contact with the ligand-expressing cell.
Additional interactions occur as AB descendants on the left and right sides of the embryo meet at the ventral midline. In wild-type embryogenesis, an AB descendant called ABprpapppp on the right side of the vental midline produces a pair of valve cells that link the intestine to the rectum. This descendant contacts a bilaterally symmetrical cell on the left side of the embryo, called ABplpapppp, that normally produces a neuron and a rectal epithelial cell (Sulston et al., 1983). However, if the right cell is killed immediately after its birth, the left cell instead produces the valve cell pair (Bowerman et al., 1992). If the left and right cells are prevented from contacting each other, both produce valve cell pairs. Because both the left and right cells produce valve cell pairs in lag-2 mutants, the Notch pathway appears to be required to restrict valve cell development to the left cell (Bowerman et al., 1992).
lin-12 mutants hatch as larvae with defects in two ventral midline cells called G2 and W; loss of function mutants appear to have two W-like cells, while gain of function mutants have two G2-like cells (Greenwald et al., 1983). Both G2 and W express LIN-12 prominently during ventral enclosure (J. Priess, unpublished); these cells do not appear to contact the MS descendants, and the ligand-expressing cells for this interaction are not known.
With the possible exception of the valve cell and G2/W interactions, all of the Notch interactions that have been characterized thus far in the embryo are inductive, in contrast to lateral interactions between initially equivalent cells that can occur during postembryonic development (see LIN-12/Notch signaling in C. elegans). Perhaps the twin constraints of rapid development and relatively few cells in early embryogenesis favors the precision of asymmetric, inductive interactions, where the outcome is invariant. Future studies should determine whether examples of lateral interactions occur in later embryogenesis.
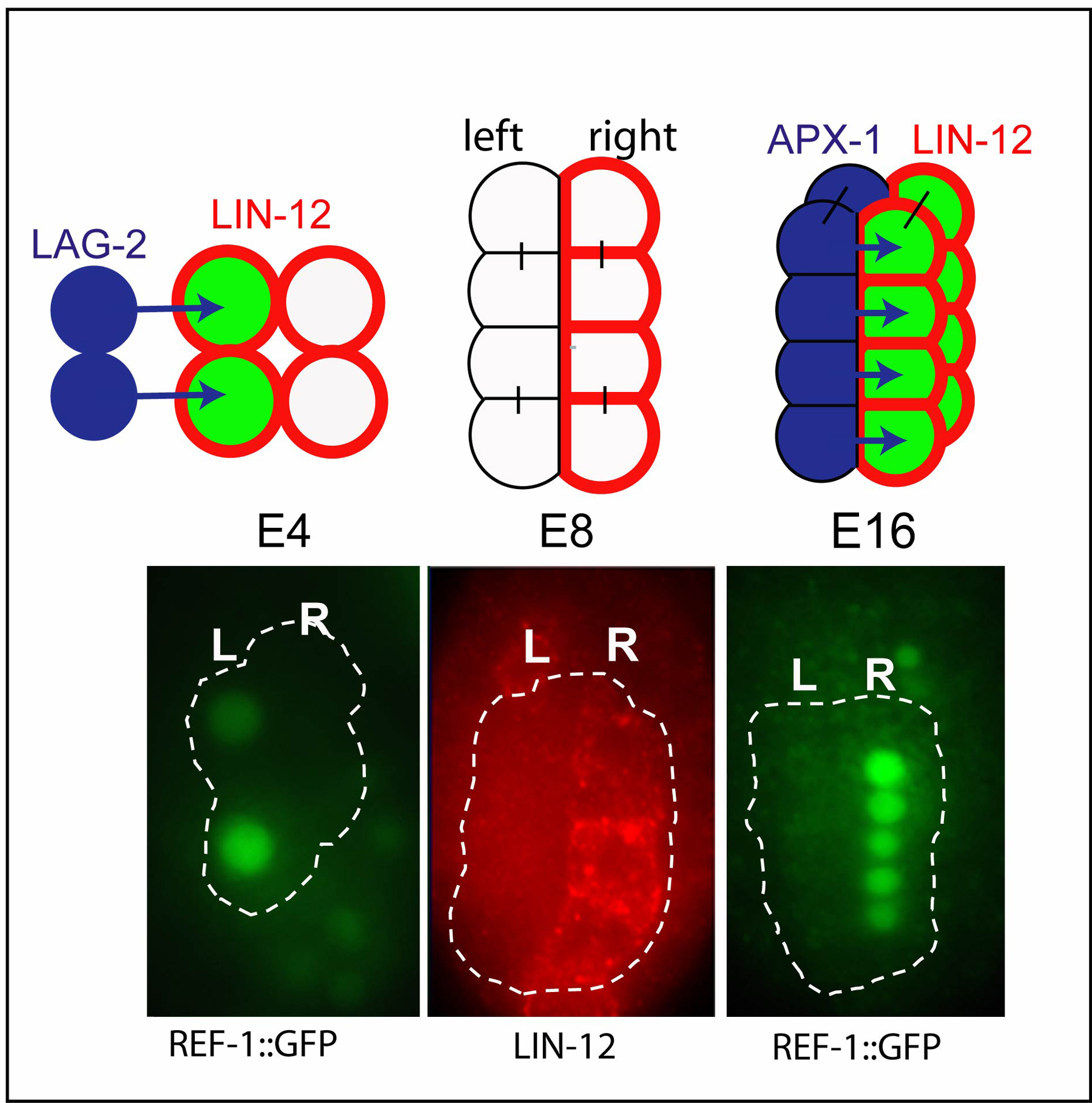
Several P1 descendants express GLP-1/Notch or LIN-12/Notch in wild-type development (J. Priess, unpublished). However, Notch interactions thus far have been described only for the E blastomere, a P1 descendant that generates the entire intestine (Hermann et al., 2000). Stages of intestinal development are designated as E2, E4, etc. to indicate the number of E descendants in the intestinal primordium; Notch interactions mediated by LIN-12 occur at both the E4 and E16 stages (Figure 8). At the E4 stage, all four E descendants express LIN-12/Notch, but only the two left cells contact LAG-2/Delta-expressing cells outside the primordium. Notch signaling activates REF-1 expression in the left E4 cells, and REF-1 functions to down-regulate LIN-12 expression in the left E8 cells (Neves and Priess, 2005). Thus LIN-12 remains only in the right E8 and early E16 cells, where it is activated at the E16 stage by the ligand APX-1/Delta (Hermann et al., 2000). The E16 interaction leads to a second wave of REF-1 expression on the right side (Figure 8; Neves and Priess, 2005). The E16 interaction initiates asymmetrical movements that eventually twist the developing intestine. Thus, in the intestinal lineage Notch signaling regulates cell behavior during morphogenesis, rather than regulating cell fate specification. How ref-1, or other possible targets of the E16 interaction, lead to intestinal twist is not yet known.
|
Figure 8. The E4 and E8 Notch interactions in the intestinal primordium are illustrated in the top row. Cell in the intestinal primordium (circled in bottom panels) express REF-1 in the left E4, leading to the down-regulation of LIN-12 at the E8 stage. The E16 interaction then induces REF-1 expression in the right cells. Adapted from Neves and Priess (2005).
As illustrated by Notch signaling in the intestinal primordium, the complex patterns of interactions during embryogenesis stem in part from Notch-mediated regulation of Notch signaling components. These links allow one interaction to pattern subsequent interactions. The activation of maternally-expressed GLP-1/Notch by the first two interactions in the AB lineage induces the embryonic expression of LIN-12/Notch (Figure 3; Moskowitz and Rothman, 1996). Thus ABp descendants, and a subset of ABa descendants, can participate in later Notch interactions. These examples of Notch-mediated induction of LIN-12 are in contrast to the intestinal interactions that down-regulate LIN-12 expression (Hermann et al., 2000). Similarly, the third Notch interaction in the AB lineage down-regulates LIN-12 expression (Neves and Priess, 2005). In addition to activating LIN-12 expression, the second Notch interaction serves to repress LAG-2/Delta expression (Moskowitz and Rothman, 1996). This interaction restricts LAG-2 to ABala descendants that eventually serve as signaling cells for the third Notch interaction. Notch-mediated repression of LAG-2 and LIN-12 requires REF-1 or REF-1 family members (Neves and Priess, 2005). The mechanism of Notch-mediated induction of LIN-12, or Notch-mediated repression of LIN-12, is not yet known.
C. elegans provides an excellent system for a detailed understanding of how one of the key metazoan signaling pathways, the Notch pathway, functions during development. Our current view of the Notch network in the embryo will undoubtedly become more elaborate as additional interactions are discovered. For example, the finding that Notch signaling functions in intestinal morphogenesis raises the possibility of other roles in tissue morphogenesis that should be examined. Notch signaling might also have a role in neuronal pathfinding in late embryogenesis, as it appears to have in Drosophila (Crowner et al., 2003). Such roles might be revealed by detailed anatomical studies of lin-12 glp-1 double mutants.
A central question remains how Notch signaling is linked to so many different cell fates or cell behaviors during embryogenesis. A ternary complex consisting of the DNA-binding protein LAG-1, the Notch intracellular domain, and SEL-8/LAG-3 can function as a strong transcriptional activator in yeast assays (Petcherski and Kimble, 2000). However, there are over 30,000 potential binding sites for LAG-1 in the C. elegans genome, making it likely that Notch activation is influenced by combinatorial factors that have not yet been identified. The REF-1 family of Notch targets appears to be the major effector of Notch-mediated repression in the embryo, and analysis of this gene family should provide insight into Notch-regulated transcription.
REF-1 proteins are distant relatives of the HES proteins that are Notch targets in other animals, indicating that a major regulatory output of Notch signaling has been conserved in evolution. Analysis of the REF-1 family in C. elegans embryos provides two examples where a Notch target also is expressed in Notch-independent cells. The REF-1 proteins may have similar functions in both Notch-activated and Notch-independent cells, such that Notch signaling effectively serves to replicate a Notch-independent pattern of development in additional parts of the embryo. In future studies, it will be interesting in to see whether this theme continues for other Notch targets in C. elegans and in other animals.
There are numerous phenotypic differences between embryos depleted of the REF-1 proteins and embryos defective in some of the early Notch interactions, suggesting that there are several additional Notch targets remaining to be discovered. Those targets may be found through genetic screens for mutants lacking Notch-dependent cell types, or through bioinformatic studies as Notch-responsive enhancer elements are analyzed in detail (Yoo et al., 2004). The well-characterized fate transformations that result from Notch signaling will provide a rigorous test for deciding whether and how candidate target genes contribute to cell fate decisions.
Alper, S., and Kenyon, C. (2001). REF-1, a protein with two bHLH domains, alters the pattern of cell fusion in C. elegans by regulating Hox protein activity. Development 128, 1793–1804. Abstract
Artavanis-Tsakonas, S., Rand, M.D., and Lake, R.J. (1999). Notch signaling: cell fate control and signal integration in development. Science 284, 770–776. Abstract Article
Austin, J., and Kimble, J. (1987). glp-1 is required in the germ line for regulation of the decision between mitosis and meiosis in C. elegans. Cell 51, 589–599. Abstract Article
Azzaria, M., Goszczynski, B., Chung, M.A., Kalb, J.M., and McGhee, J.D. (1996). A fork head/HNF-3 homolog expressed in the pharynx and intestine of the Caenorhabditis elegans embryo. Dev. Biol. 178, 289–303. Abstract Article
Berset, T., Hoier, E.F., Battu, G., Canevascini, S., and Hajnal, A. (2001). Notch inhibition of RAS signaling through MAP kinase phosphatase LIP-1 during C. elegans vulval development. Science 291, 1055–1058. Abstract Article
Bowerman, B., Eaton, B.A., and Priess, J.R. (1992). skn-1, a maternally expressed gene required to specify the fate of ventral blastomeres in the early C. elegans embryo. Cell 68, 1061–1075. Abstract Article
Bowerman, B., Tax, F.E., Thomas, J.H., and Priess, J.R. (1992). Cell interactions involved in development of the bilaterally symmetrical intestinal valve cells during embryogenesis in Caenorhabditis elegans. Development 116, 1113–1122. Abstract
Broitman-Maduro, G., Maduro, M.F., and Rothman, J.H. (2005). The noncanonical binding site of the MED-1 GATA factor defines differentially regulated target genes in the C. elegans mesendoderm. Dev. Cell 8, 427–433. Abstract Article
Crowner, D., Le Gall, M., Gates, M.A., and Giniger, E. (2003). Notch steers Drosophila ISNb motor axons by regulating the Abl signaling pathway. Curr. Biol. 13, 967–972. Abstract Article
Doyle, T.G., Wen, C., and Greenwald, I. (2000). SEL-8, a nuclear protein required for LIN-12 and GLP-1 signaling in Caenorhabditis elegans. Proc. Natl. Acad. Sci. USA 97, 7877–7881. Abstract Article
Edgar, L.G., Carr, S., Wang, H., and Wood, W.B. (2001). Zygotic expression of the caudal homolog pal-1 is required for posterior patterning in Caenorhabditis elegans embryogenesis. Dev. Biol. 229, 71–88. Abstract Article
Evans, T.C., Crittenden, S.L., Kodoyianni, V., and Kimble, J. (1994). Translational control of maternal glp-1 mRNA establishes an asymmetry in the C. elegans embryo. Cell 77, 183–194. Abstract Article
Finney, M., and Ruvkun, G. (1990). The unc-86 gene product couples cell lineage and cell identity in C. elegans. Cell 63, 895–905. Abstract Article
Gendreau, S.B., Moskowitz, I.P., Terns, R.M., and Rothman, J.H. (1994). The potential to differentiate epidermis is unequally distributed in the AB lineage during early embryonic development in C. elegans. Dev. Biol. 166, 770–781. Abstract Article
Good, K., Ciosk, R., Nance, J., Neves, A., Hill, R.J., and Priess, J.R. (2004). The T-box transcription factors TBX-37 and TBX-38 link GLP-1/Notch signaling to mesoderm induction in C. elegans embryos. Development 131, 1967–1978. Abstract Article
Goutte, C., Hepler, W., Mickey, K.M., and Priess, J.R. (2000). aph-2 encodes a novel extracellular protein required for GLP-1-mediated signaling. Development 127, 2481–2492. Abstract
Goutte, C., Tsunozaki, M., Hale, V.A., and Priess, J.R. (2002). APH-1 is a multipass membrane protein essential for the Notch signaling pathway in Caenorhabditis elegans embryos. Proc. Natl. Acad. Sci. USA 99, 775–779. Abstract Article
Greenwald, I.S., Sternberg, P.W., and Horvitz, H.R. (1983). The lin-12 locus specifies cell fates in Caenorhabditis elegans. Cell 34, 435–444. Abstract Article
Gupta, B.P., and Sternberg, P.W. (2002). Tissue-specific regulation of the LIM homeobox gene lin-11 during development of the Caenorhabditis elegans egg-laying system. Dev. Biol. 247, 102–115. Abstract Article
Hermann, G.J., Leung, B., and Priess, J.R. (2000). Left-right asymmetry in C. elegans intestine organogenesis involves a LIN-12/Notch signaling pathway. Development 127, 3429–3240. Abstract
Horner, M.A., Quintin, S., Domeier, M.E., Kimble, J., Labouesse, M., and Mango, S.E. (1998). pha-4, an HNF-3 homolog, specifies pharyngeal organ identity in Caenorhabditis elegans. Genes Dev. 12, 1947–1952. Abstract
Hutter, H., and Schnabel, R. (1994). glp-1 and inductions establishing embryonic axes in C. elegans. Development 120, 2051–2064. Abstract
Hutter, H., and Schnabel, R. (1995a). Establishment of left-right asymmetry in the Caenorhabditis elegans embryo: a multistep process involving a series of inductive events. Development 121, 3417–3424. Abstract
Hutter, H., and Schnabel, R. (1995b). Specification of anterior-posterior differences within the AB lineage in the C. elegans embryo: a polarising induction. Development 121, 1559–1568. Abstract
Kalb, J.M., Lau, K.K., Goszczynski, B., Fukushige, T., Moons, D., Okkema, P.G., and McGhee, J.D. (1998). pha-4 is Ce-fkh-1, a fork head/HNF-3α,β,γ homolog that functions in organogenesis of the C. elegans pharynx. Development 125, 2171–2180. Abstract
Kaletta, T., Schnabel, H., and Schnabel, R. (1997). Binary specification of the embryonic lineage in Caenorhabditis elegans. Nature 390, 294–298. Abstract Article
Lambie, E.J., and Kimble, J. (1991). Two homologous regulatory genes, lin-12 and glp-1, have overlapping functions. Development 112, 231–240. Abstract
Lamont, L.B., Crittenden, S.L., Bernstein, D., Wickens, M., and Kimble, J. (2004). FBF-1 and FBF-2 regulate the size of the mitotic region in the C. elegans germline. Dev. Cell 7, 697–707. Abstract Article
Lin, R., Hill, R.J., and Priess, J.R. (1998). POP-1 and anterior-posterior fate decisions in C. elegans embryos. Cell 92, 229–239. Abstract Article
Lin, R., Thompson, S., and Priess, J.R. (1995). pop-1 encodes an HMG box protein required for the specification of a mesoderm precursor in early C. elegans embryos. Cell 83, 599–609. Abstract Article
Lo, M.C., Gay, F., Odom, R., Shi, Y., and Lin, R. (2004). Phosphorylation by the β-catenin/MAPK complex promotes 14-3-3-mediated nuclear export of TCF/POP-1 in signal-responsive cells in C. elegans. Cell 117, 95–106. Abstract Article
Maduro, M.F., Lin, R., and Rothman, J.H. (2002). Dynamics of a developmental switch: recursive intracellular and intranuclear redistribution of Caenorhabditis elegans POP-1 parallels Wnt-inhibited transcriptional repression. Dev. Biol. 248, 128–142. Abstract Article
Maduro, M.F., Meneghini, M.D., Bowerman, B., Broitman-Maduro, G., and Rothman, J.H. (2001). Restriction of mesendoderm to a single blastomere by the combined action of SKN-1 and a GSK-3β homolog is mediated by MED-1 and -2 in C. elegans. Mol. Cell 7, 475–485. Abstract Article
Mango, S.E., Lambie, E.J., and Kimble, J. (1994). The pha-4 gene is required to generate the pharyngeal primordium of Caenorhabditis elegans. Development 120, 3019–3031. Abstract
Mango, S.E., Thorpe, C.J., Martin, P.R., Chamberlain, S.H., and Bowerman, B. (1994). Two maternal genes, apx-1 and pie-1, are required to distinguish the fates of equivalent blastomeres in the early Caenorhabditis elegans embryo. Development 120, 2305–2315.
Mello, C.C., Draper, B.W., Krause, M., Weintraub, H., and Priess, J.R. (1992). The pie-1 and mex-1 genes and maternal control of blastomere identity in early C. elegans embryos. Cell 70, 163–176. Abstract Article
Mello, C.C., Draper, B.W., and Priess, J.R. (1994). The maternal genes apx-1 and glp-1 and establishment of dorsal-ventral polarity in the early C. elegans embryo. Cell 77, 95–106. Abstract Article
Mickey, K.M., Mello, C.C., Montgomery, M.K., Fire, A., and Priess, J.R. (1996). An inductive interaction in 4-cell stage C. elegans embryos involves APX-1 expression in the signalling cell. Development 122, 1791–1798. Abstract
Moskowitz, I.P., Gendreau, S.B., and Rothman, J.H. (1994). Combinatorial specification of blastomere identity by glp-1-dependent cellular interactions in the nematode Caenorhabditis elegans. Development 120, 3325–3338. Abstract
Moskowitz, I.P., and Rothman, J.H. (1996). lin-12 and glp-1 are required zygotically for early embryonic cellular interactions and are regulated by maternal GLP-1 signaling in Caenorhabditis elegans. Development 122, 4105–4117. Abstract
Neves, A., and Priess, J.R. (2005). The REF-1 family of bHLH transcription factors pattern C. elegans embryos through Notch-dependent and Notch-independent pathways. Dev. Cell. 8, 867–879. Abstract Article
Park, F.D., and Priess, J.R. (2003). Establishment of POP-1 asymmetry in early C. elegans embryos. Development 130, 3547–3556. Abstract Article
Park, F.D., Tenlen, J.R., and Priess, J.R. (2004). C. elegans MOM-5/frizzled functions in MOM-2/Wnt-independent cell polarity and is localized asymmetrically prior to cell division. Curr. Biol. 14, 2252–2258. Abstract Article
Petcherski, A.G., and Kimble, J. (2000). LAG-3 is a putative transcriptional activator in the C. elegans Notch pathway. Nature 405, 364–368. Abstract Article
Priess, J.R., Schnabel, H., and Schnabel, R. (1987). The glp-1 locus and cellular interactions in early C. elegans embryos. Cell 51, 601–611. Abstract Article
Priess, J.R., and Thomson, J.N. (1987). Cellular interactions in early C. elegans embryos. Cell 48, 241–250. Abstract Article
Robertson, S.M., Shetty, P., and Lin, R. (2004). Identification of lineage-specific zygotic transcripts in early Caenorhabditis elegans embryos. Dev. Biol. 276, 493–507. Abstract Article
Rocheleau, C.E., Downs, W.D., Lin, R., Wittmann, C., Bei, Y., Cha, Y.H., Ali, M., Priess, J.R., and Mello, C.C. (1997). Wnt signaling and an APC-related gene specify endoderm in early C. elegans embryos. Cell 90, 707–716. Abstract Article
Rocheleau, C.E., Yasuda, J., Shin, T.H., Lin, R., Sawa, H., Okano, H., Priess, J.R., Davis, R.J., and Mello, C.C. (1999). WRM-1 activates the LIT-1 protein kinase to transduce anterior/posterior polarity signals in C. elegans. Cell 97, 717–726. Abstract Article
Shelton, C.A., and Bowerman, B. (1996). Time-dependent responses to glp-1-mediated inductions in early C. elegans embryos. Development 122, 2043–2050. Abstract
Strutt, D.I. (2001). Asymmetric localization of frizzled and the establishment of cell polarity in the Drosophila wing. Mol. Cell 7, 367–375. Abstract Article
Sulston, J.E., Schierenberg, E., White, J.G., and Thomson, J.N. (1983). The embryonic cell lineage of the nematode Caenorhabditis elegans. Dev. Biol. 100, 64–119. Abstract Article
Thorpe, C.J., Schlesinger, A., Carter, J.C., and Bowerman, B. (1997). Wnt signaling polarizes an early C. elegans blastomere to distinguish endoderm from mesoderm. Cell 90, 695–705. Abstract Article
Way, J.C., Wang, L., Run, J.Q., and Hung, M.S. (1994). Cell polarity and the mechanism of asymmetric cell division. Bioessays 16, 925–931. Abstract Article
Wilkinson, H.A., Fitzgerald, K., and Greenwald, I. (1994). Reciprocal changes in expression of the receptor lin-12 and its ligand lag-2 prior to commitment in a C. elegans cell fate decision. Cell 79, 1187–1198. Abstract Article
Wittmann, C., Bossinger, O., Goldstein, B., Fleischmann, M., Kohler, R., Brunschwig, K., Tobler, H., and Muller, F. (1997). The expression of the C. elegans labial-like Hox gene ceh-13 during early embryogenesis relies on cell fate and on anteroposterior cell polarity. Development 124, 4193–4200. Abstract
*Edited by Geraldine Seydoux. Last revised August 4, 2005. Published June 25, 2005. This chapter should be cited as: Priess, J. Notch signaling in the C. elegans embryo (June 25, 2005), WormBook, ed. The C. elegans Research Community, WormBook, doi/10.1895/wormbook.1.4.1, http://www.wormbook.org.
Copyright: © 2005 James R. Priess. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
§To whom correspondence should be addressed. E-mail: [email protected]
 All WormBook content, except where otherwise noted, is licensed under a Creative Commons Attribution License.
All WormBook content, except where otherwise noted, is licensed under a Creative Commons Attribution License.