Embryo series courtesy of Einhard Schierenberg
Embryo series courtesy of Einhard SchierenbergTable of Contents
Abstract
Sexual reproduction requires the production of haploid gametes (sperm and egg) with only one copy of each chromosome; fertilization then restores the diploid chromosome content in the next generation. This reduction in genetic content is accomplished during a specialized cell division called meiosis, in which two rounds of chromosome segregation follow a single round of DNA replication. In preparation for the first meiotic division, homologous chromosomes pair and synapse, creating a context that promotes formation of crossover recombination events. These crossovers, in conjunction with sister chromatid cohesion, serve to connect the two homologs and facilitate their segregation to opposite poles during the first meiotic division. During the second meiotic division, which is similar to mitosis, sister chromatids separate; the resultant products are haploid cells that become gametes.
In Caenorhabditis elegans (and most other eukaryotes) homologous pairing and recombination are required for proper chromosome inheritance during meiosis; accordingly, the events of meiosis are tightly coordinated to ensure the proper execution of these events. In this chapter, we review the seminal events of meiosis: pairing of homologous chromosomes, the changes in chromosome structure that chromosomes undergo during meiosis, the events of meiotic recombination, the differentiation of homologous chromosome pairs into structures optimized for proper chromosome segregation at Meiosis I, and the ultimate segregation of chromosomes during the meiotic divisions. We also review the regulatory processes that ensure the coordinated execution of these meiotic events during prophase I.
CO–crossover, DSB–double strand break, HR–homologous recombination, HRR–homology recognition region, IR–ionizing radiation, NCO–non-crossover, NHEJ–non-homologous end joining, PC–pairing center, SC–synaptonemal complex
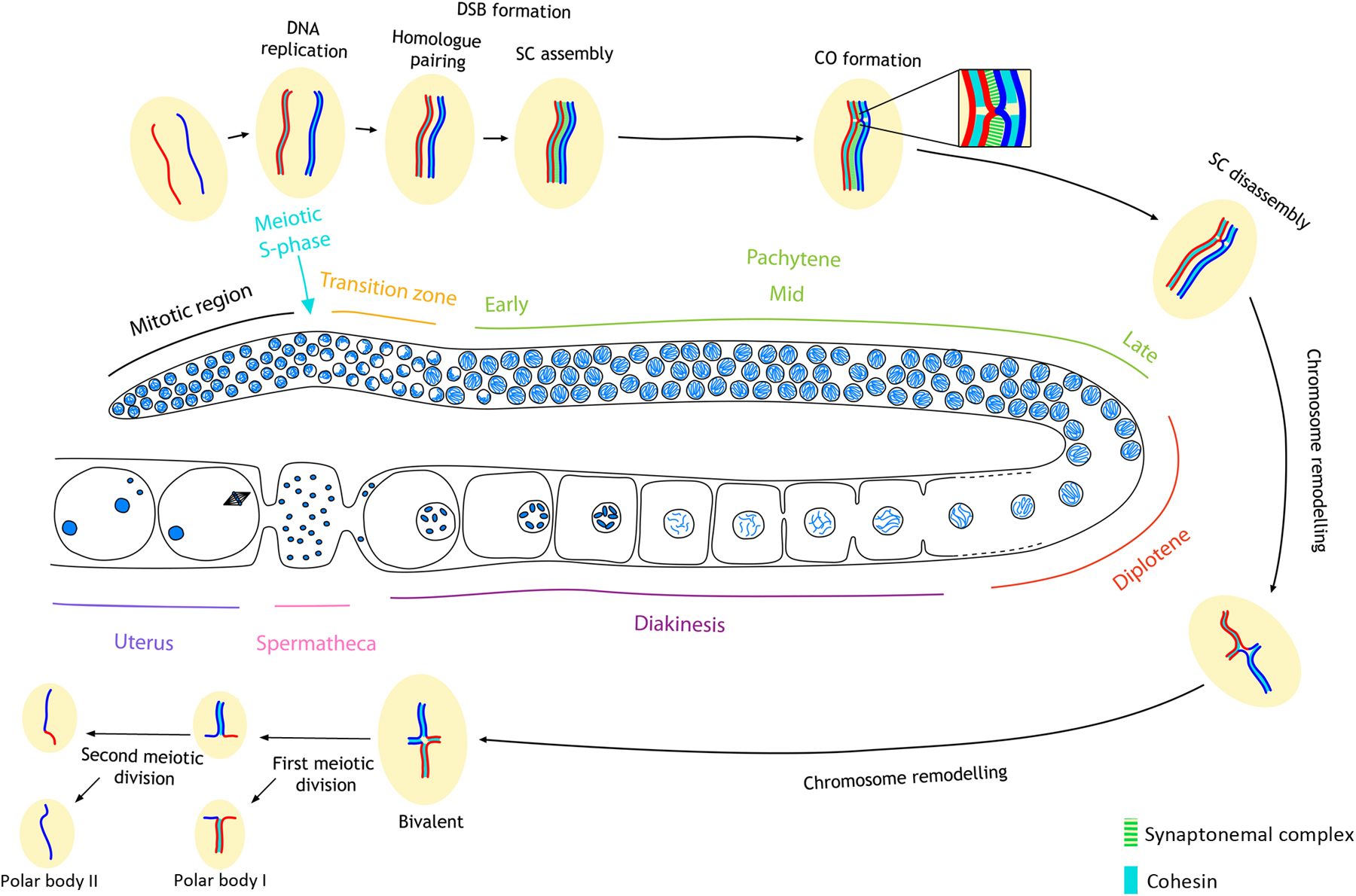
Sexual reproduction requires the generation of haploid gametes from diploid precursors through the specialized cell division program of meiosis. This reduction in ploidy is essential to ensure the restoration of diploidy upon fertilization and requires completion of several key events (Figure 1). During early prophase (leptotene and zygotene stages), each chromosome must locate and recognize its appropriate homologous pairing partner and align with it. During the zygotene stage, a specialized protein structure called the synaptonemal complex (SC) assembles between the aligned chromosomes to hold homologs together; full synapsis of homologs defines the pachytene stage. Crossover (CO) recombination events must be completed between the DNA molecules of the aligned and synapsed homologs, a process started by the deliberate formation of DNA double strand breaks (DSBs). Crossing over is essential for the formation of chiasmata, connections between homologs that become evident upon structural remodeling of chromosomes during later stages of meiotic prophase (diplotene and diakinesis). Therefore, there are multiple surveillance mechanisms that act to ensure that each homolog pair undergoes an exchange. Late prophase remodeling of chromosome pairs connected by chiasmata results in bivalents wherein the connected homologs are oriented away from each other; this promotes bipolar attachment of homologs to the meiosis I spindle, leading to segregation of homologous chromosomes at anaphase I. The separation of sister chromatids on the meiosis II spindle completes the meiotic program.
|
Figure 1: Diagram of meiotic events during oogenesis in the C. elegans germ line. For simplicity, a single pair of homologous chromosomes is shown. DNA replication occurs at the onset of meiosis. During the transition zone in early meiotic prophase (leptotene and zygotene), homolog pairing is achieved and DSBs are formed. By early pachytene, SC assembly is completed, and by late pachytene, the process of homologous recombination leads to the formation of inter-homolog CO events. Following SC disassembly during late prophase, homologous chromosomes remain linked by chiasmata, physical attachments provided by COs in combination with sister chromatid cohesion. In the first meiotic division parental homologs are disjoined to reduce ploidy to the haploid state, and in the second division sister chromatids are disjoined. Recombination leads to genetic exchange as indicated by the different coloring of the chromosomes. Note: during oocyte meiosis, one of the two end products of each division gets extruded as a polar body.
Despite the fundamental importance of meiosis in sexual reproduction, many basic questions about the process and the underlying mechanisms remain unanswered. Over the past twenty years, Caenorhabditis elegans has emerged as a major model organism for investigating meiotic mechanisms. Several features of C. elegans biology have contributed to this emergence. The worm germ line is especially amenable to high-resolution cytological analysis of chromosome and nuclear organization in the context of whole mount preparations that preserve 3D nuclear architecture. Importantly, each germ line contains a complete time course of meiosis, with nuclei organized in a temporal/spatial gradient corresponding to the stages of meiotic prophase (Figure 1 and Figure 2). Further, the chromosomal basis of sex determination can be exploited to identify meiotic mutants on the basis of sex chromosome missegregation. The availability of worms expressing GFP::histone has also made it possible to screen for mutants based on lack of chiasmata connecting homologs at the end of meiotic prophase. Mutant hermaphrodites can still produce a few percent euploid survivors even if all six chromosome pairs lack chiasmata, a feature that has greatly facilitated analysis of meiotic mutants. Germline mRNA expression profiles have accelerated identification of molecular defects associated with meiotic mutants and have provided a basis for identification of candidate genes tested for meiotic roles in targeted RNAi screens. Finally, C. elegans has a robust tradition of investigating the genetic behavior of chromosome rearrangements, which has led to the discovery of cis-acting chromosome features that govern meiotic chromosome behavior.
|
Figure 2: The hermaphrodite germ line contains a complete time course of meiotic prophase. Projection of a dissected, three-dimensionally intact hermaphrodite germ line stained with DAPI to visualize chromatin. Stages of meiotic prophase and magnification of a representative nucleus of each meiotic prophase stage are depicted below the germ line. White lines indicate boundaries between germline regions containing nuclei at indicated stages of meiotic prophase. Left end of the germ line contains a compartment of mitotically proliferating cells followed by nuclei undergoing meiotic DNA replication (meiotic entrance). The transition zone contains nuclei in leptotene/zygotene, with chromatin tightly clustered in one half of the nucleus, giving a characteristic half-moon shape appearance. As nuclei move from the transition zone into the pachytene region, paired and aligned homologous chromosomes can be seen distributed around the periphery of each nucleus. Diplotene is characterized by chromatin condensation; at this stage, bivalents (pairs of homologous of chromosomes attached by chiasmata) become visible as 6 separate chromatin structures. Chromosome condensation continues during diakinesis, when the 6 bivalents can be easily visualized within the nucleus. Nuclei move down the gonad tube, transitioning between the stages described above with a speed of approximately one cell row per hour. Inset nuclei from the various stages of prophase I are shown to scale with each other.
This chapter begins with a “parts list” of meiotic machinery components identified in C. elegans, followed by a description of the events of meiosis that integrates information about the roles of these components. We will focus on oocyte meiosis, as later stages of prophase in oocytes are cytologically more accessible than during spermatocyte meiosis. We will discuss the interrelated processes of chromosome movement and pairing (Section 3), and the protein complexes that drive the dramatic changes seen in chromosome structure during meiosis (Section 4; also see Germline chromatin). We will then discuss the process of meiotic recombination (Section 5). Following chiasma formation, late pachytene bivalents differentiate around the site of the chiasma in preparation for subsequent segregation (Section 6). We round off the chapter with an overview of surveillance mechanisms that monitor meiotic events for proper completion (Section 7) and a description of the events that occur during meiotic chromosome segregation (Section 8).
Pairing of homologous chromosomes, which occurs during the leptotene and zygotene stages of prophase I (transition zone of the germ line, Figure 1), is a crucial event in meiosis. During this highly dynamic process, the two homologous copies of each chromosome find each other within the nucleus through an active search process that enables chromosomes to distinguish “self versus non-self” and assume a side-by-side alignment. This pairing is a necessary prerequisite for CO formation, and thus successful completion of meiosis. In many organisms pairing is mediated by tethering chromosome ends to the nuclear periphery where they become attached to cytoplasmic motor proteins via SUN/KASH domain protein complexes that span the nuclear envelope. The motor proteins then drive chromosome movement that is essential for the timely completion of pairing. In C. elegans cis-acting sequences near one end of each chromosome, rather than the telomere itself, assemble a nucleoprotein complex that tethers the chromosome ends to the nuclear envelope. These events occur in the transition zone and coincide with chromatin adopting a special configuration, with chromosomes pushed to one side of the nucleus opposite the nucleolus, giving the chromatin a half-moon shape (Figure 2). Pairing is complete by exit from the transition zone (MacQueen and Villeneuve, 2001). Mutations that disrupt chromosome movements also result in loss of this nuclear reorganization during leptotene/zygotene (MacQueen and Villeneuve, 2001; Couteau et al., 2004; Couteau and Zetka, 2005; Martinez-Perez and Villeneuve, 2005; Penkner et al., 2007b). The process of pairing is normally coupled with SC assembly between homologs (Section 4). However, the chromosome alignment process is genetically separable from synapsis, although both take place along the entire length of the chromosome (Pasierbek et al., 2001; MacQueen et al., 2002; Nabeshima et al., 2011).
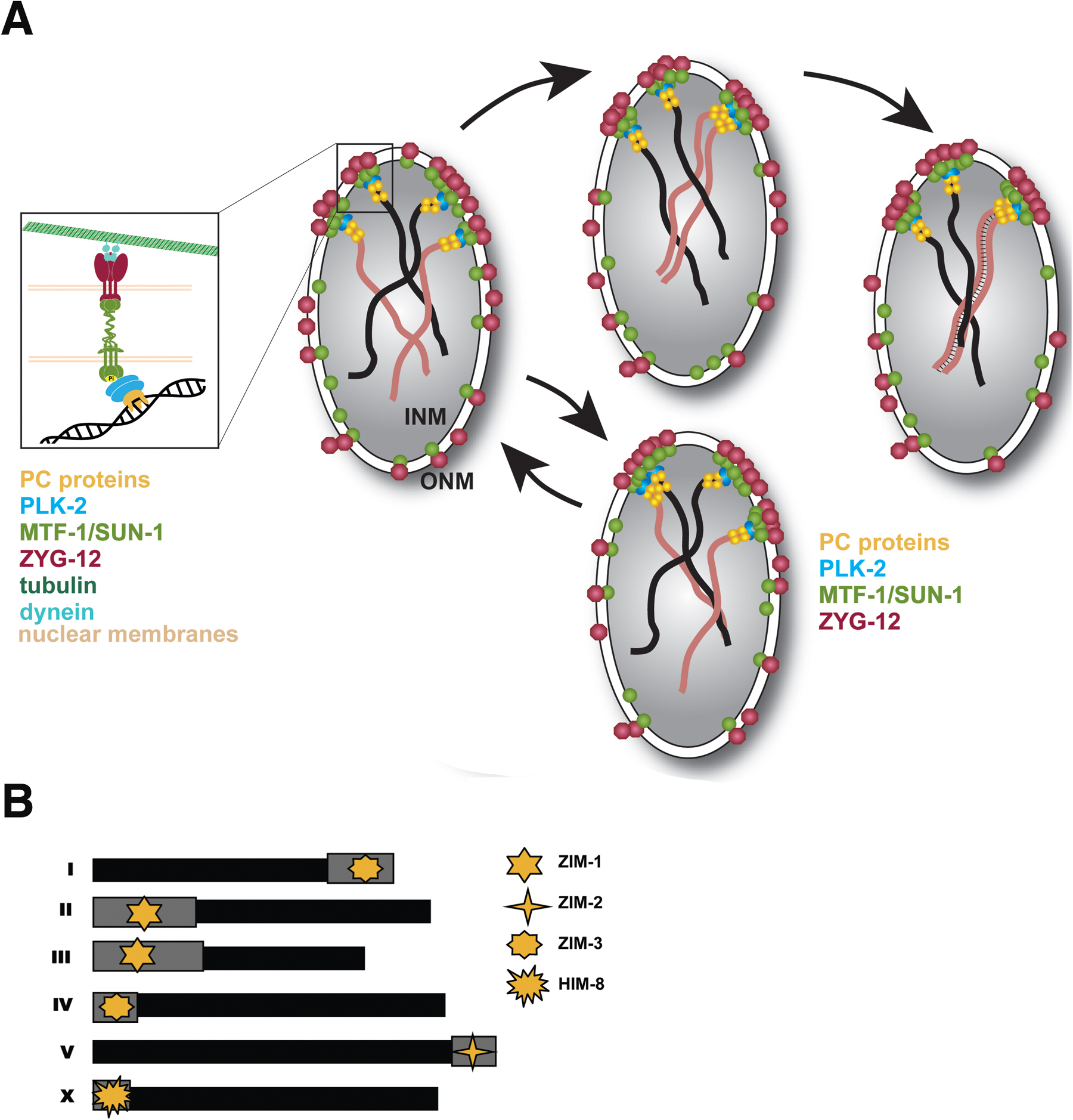
Each C. elegans chromosome has a localized cis-acting region near one end that plays crucial roles in meiotic chromosome behavior. These regions have been termed HRRs (homology recognition regions) or PCs (pairing centers) (Rosenbluth and Baillie, 1981; Rose et al., 1984; McKim et al., 1988; Herman and Kari, 1989; Villeneuve, 1994). These HRRs/PCs (hereafter, PCs) are each comprised of repetitive DNA sequences (Sanford and Perry, 2001; Phillips et al., 2009). The PC ends of chromosomes are in close proximity to the nuclear envelope in the transition zone, when the chromosome pairing process is initiated (Figure 3) (Goldstein, 1982; MacQueen et al., 2005). Chromosomes lacking PCs display severe defects in pairing and synapsis (MacQueen et al., 2005).
|
Figure 3: Chromosome movements and homolog pairing during early prophase I. (A). Model depicting the attachment of chromosomes to the nuclear envelope and their movement in transition zone nuclei. Model shows two pairs of homologous chromosomes (one pair depicted in black and a second pair depicted in brown) with their PC-bearing region associated with the nuclear envelope. See key on figure for molecular components. Inset on the left hand side of the panel shows a detailed model of the attachment site of a single chromosome. When chromosomes enter the transition zone they are connected to the nuclear envelope via interactions between PC-associated proteins (PC proteins) and a nuclear envelope-spanning SUN/KASH protein complex. Microtubule-mediated forces in the cytoplasm result in movement of these SUN/KASH complexes, causing movement of attached chromosome ends. Chromosome ends move vigorously, and tend to come together into local clusters (presumably to allow homology between adjacent chromosomes to be assessed). When homologs have identified each other and initiated pairing, the SC (depicted as thin black lines connecting the homologs) is established. Both synapsed and unsynapsed chromosomes continue to move until all chromosomes are synapsed. (B) Diagram showing the position of the PC region (grey boxes) on each chromosome and the PC-binding proteins that load to each chromosome.
In C. elegans meiosis, each PC is bound by one of four C2H2 zinc finger proteins (HIM-8, ZIM-1, ZIM-2, ZIM-3) (Figure 3). The genes encoding the HIM/ZIM proteins are found within a single operon, ensuring their coordinated germline expression (Phillips et al., 2005; Phillips and Dernburg, 2006). PC repeats are sufficient to recruit ZIM proteins as shown by the injection of plasmids carrying synthetic repeats of the PC motifs (Sanford and Perry, 2001; Phillips et al., 2009). Furthermore, PC proteins are required both for aligning homologs and for homologous synapsis of the specific chromosomes to which they are bound (Phillips et al., 2005; Phillips and Dernburg, 2006; Phillips et al., 2009; Harper et al., 2011; Labella et al., 2011). HIM-8 appears to have an additional role(s) as it is required for elongation of the X chromosomes in transition zone nuclei, where all chromosomes start to occupy extended territories, a process that may facilitate their lengthwise alignment (Nabeshima et al., 2011).
Although functionally similar, the X and autosome PCs differ with respect to the persistence and regulation of nuclear envelope attachment and association of their PC-binding proteins. HIM-8 localizes to X chromosome PCs from premeiotic stages through late pachytene (Phillips et al., 2005). Autosomal PC proteins, by contrast, are detected at PCs primarily from leptotene to early pachytene (ZIM-1 on chromosomes II and III, ZIM-2 on chromosome V and ZIM-3 on chromosomes I and IV). In contrast to HIM-8, concentration of the autosomal PC proteins at PCs depends on the CHK-2 protein kinase (Phillips et al., 2005; Phillips and Dernburg, 2006), a master regulator of early events of prophase (MacQueen and Villeneuve, 2001).
The PC nucleoprotein complexes act as recruitment sites for polo kinase, PLK-2 (or PLK-1, if PLK-2 is absent). PLK-2 induces structural reorganization of the nuclear envelope (Harper et al., 2011; Labella et al., 2011): the inner and outer nuclear envelope proteins SUN-1 and ZYG-12 relocate into pronounced aggregates corresponding to the sites where PCs localize to the nuclear envelope (Figure 3) (Penkner et al., 2009; Sato et al., 2009). SUN-1 and ZYG-12 form a functional SUN/KASH protein–protein interaction module, broadly conserved among eukaryotes (Malone et al., 2003; Fridkin et al., 2004; Penkner et al., 2007b; Minn et al., 2009), that spans the nuclear membranes and connects chromosomes to the cytoskeleton.
If the SUN/KASH interaction is abrogated, ZYG-12 retention at the outer nuclear envelope is lost (Malone et al., 2003; Penkner et al., 2007b) despite the assembly of SUN-1 aggregates with PCs at the inner nuclear envelope (Penkner et al., 2009). Therefore, the trigger for SUN-1 aggregation is transmitted from the nucleus to the cytoplasm where ZYG-12 mirrors SUN-1 aggregates. SUN-1 aggregate formation is independent of DSBs, recombination, pairing, and synapsis, but requires CHK-2 (Penkner et al., 2009; Sato et al., 2009). SUN-1/ZYG-12 aggregates at autosomal PC attachment sites are found in leptotene/zygotene (TZ). In contrast the SUN-1/ZYG-12 aggregates around the X chromosome PC persist throughout early pachytene. In mid- and late pachytene, most nuclei lack SUN-1/ZYG-12 aggregates, despite the presence of a HIM-8 focus at the nuclear envelope (Phillips et al., 2005; Penkner et al., 2009; Sato et al., 2009). Disappearance of SUN-1/ZYG-12 aggregates correlates with the establishment of full synapsis and relocalization of PLK-2 from PCs to the SC (Harper et al., 2011; Labella et al., 2011). The recruitment of PLK-2 to PCs and the subsequent formation of dynamic SUN-1/ZYG-12 aggregates is essential to ensure faithful SC assembly, as mutants defective in sun-1, zyg-12, plk-2, or him-8/zimΔ (lacking all PC-binding proteins) display aberrant synapsis (Penkner et al., 2007b; Sato et al., 2009; Harper et al., 2011; Labella et al., 2011; Woglar et al., 2013).
PCs play a prominent role in promoting homolog recognition, but several lines of evidence suggest that they cannot be the sole determinants of chromosome identity. Some PC proteins (ZIM-1 and ZIM-3) localize to more than one chromosome; despite this, nonhomologous pairing between different ZIM-1 or ZIM-3-binding PCs is not detected in wild-type animals (Phillips et al., 2009). Moreover, pre-synaptic alignment has been demonstrated along the entire length of chromosomes (Nabeshima et al., 2011). The chromodomain protein MRG-1, present on the autosomes, plays a role in the non-PC mediated homolog alignment and its absence leads to defects in homolog alignment and synapsis of non-PC regions (Dombecki et al., 2011). Mutations in the gene encoding the serine/threonine phosphatase PPH-4.1 cause defective autosomal chromosome pairing and synapsis between non-homologous chromosomes. The relevant targets for PPH-4.1 are still unknown (Sato-Carlton et al., 2014). In C. elegans, homologous recombination is not required for establishment of homolog alignment since spo-11 mutants, in which meiotic DSBs are not formed, are proficient to pair and synapse with their homologs (Dernburg et al., 1998).
PCs display a highly dynamic behavior during leptotene/zygotene (Penkner et al., 2009; Sato et al., 2009; Baudrimont et al., 2010); in contrast, other chromosomal regions are relatively static (Wynne et al., 2012). During leptotene/zygotene, PCs have a strong tendency to come together into transient local clusters. From there they may continue to move in groups/pairs or dissociate and resume independent movement (Figure 3). Tracking PC movements showed that they comprise both small short-range tracks and long saltatory trajectories (up to 2 µm) with an average track length of 0.5 µm and an average speed of 0.19 µm/sec (Penkner et al., 2009; Sato et al., 2009; Baudrimont et al., 2010; Wynne et al., 2012). Chromosomes progressively pair and synapse while progressing through leptotene/zygotene. In spite of this, the characteristics of chromosome movement remain essentially the same (Baudrimont et al., 2010; Wynne et al., 2012), suggesting that PC-mediated chromosome end movement does not cease once synapsis of a given chromosome pair is achieved.
Chromosome movement in the gonad relies on microtubules (Sato et al., 2009; Wynne et al., 2012). Further, ZYG-12 recruits components of the dynein motor complex to the cytoplasmic side of the PC attachments (Sato et al., 2009; Labrador et al., 2013) to mobilize chromosome ends in leptotene/zygotene and to facilitate pairing and synapsis. Dynein knock-down, ATP depletion, or a specific allele affecting the mitochondria-localized SPD-3 protein (with a likely role in energy production for dynein function) consistently result in reduced pairing and in synapsis with non-homologs (Labrador et al., 2013). However, a direct role of dynein-driven chromosome movements in licensing SC assembly has also been proposed, since severe dynein knockdown inhibited SC assembly (Sato et al., 2009). Sato et al. (2009) further propose that dynein functions to oppose inappropriate chromosome interactions, enabling dissociation of non-homologous chromosomes.
Upon completion of pairing and synapsis, meiocytes enter early pachytene and chromosome clustering is loosened but not completely abrogated. SUN-1/ZYG-12 aggregates dissolve around the autosome PCs, whereas they persist longer at the X chromosome PCs, which remain mobile in early pachytene (Penkner et al., 2009; Sato et al., 2009; Wynne et al., 2012). The study of the him-19 mutant revealed that regulation of chromosome end mobilization deteriorates with progressive age in this mutant (Penkner et al., 2009; Tang et al., 2010). The decreased movement and associated increase in nondisjunction with age might suggest an underlying cause for age-related chromosomal abnormalities in systems where meiosis continues throughout one's lifetime, such as during human spermatogenesis.
The meiotic program involves dramatic changes in chromosome structure, which are driven by the regulated association and dissociation of different protein complexes from chromosomes. Some of these protein complexes represent meiosis-specific elaborations upon structures that occur in mitotically cycling cells. Critically important is the loading of multiple, partially redundant cohesin complexes (Section 4.1), which tether sister chromatids together and serve as the basis for assembly of axial elements containing four HORMA-domain proteins (Section 4.2). During pachytene, the axial elements of paired homologous chromosomes are connected by yet another meiosis-specific structure, the SC (Section 4.3). Proper assembly of the SC depends on axis assembly, and in turn is required for regulated formation of meiotic COs between homologs (and thus, proper chromosome segregation) (Section 5 and Section 6). The correct execution of these structural changes is essential to promote pairing, CO formation, and chromosome segregation.
The acquisition of meiosis-specific chromosome features starts with DNA replication, during which sister chromatid cohesion must be established. Meiotic S-phase is twice as long as that of cells in the mitotic compartment of the germ line (Jaramillo-Lambert et al., 2007), perhaps reflecting the time required to adopt the unique chromosome configurations required for meiotic success. C. elegans expresses three different meiosis-specific cohesin complexes differing in their kleisin subunit: REC-8, and the nearly identical and functionally redundant COH-3 and COH-4 (Pasierbek et al., 2001; Severson et al., 2009; Severson and Meyer, 2014). REC-8 is present during meiotic DNA replication—it provides cohesion independently of DSBs, and its loading onto chromosomes depends on the HORMA-domain protein HTP-3, and on TIM-1 (ortholog of the TIMELESS clock protein) (Pasierbek et al., 2001; Chan et al., 2003; Severson et al., 2009; Severson and Meyer, 2014). In contrast, COH-3/4 complexes are only detected in meiotic nuclei following the completion of S-phase, their ability to provide cohesion requires DSBs, and their loading is independent of HTP-3 and TIM-1 (Severson and Meyer, 2014). Loss of cohesion, evidenced by extensive separation of sister chromatids in diakinesis oocytes, is observed in mutants lacking REC-8, COH-3, and COH-4, but not in rec-8 single or coh-3 coh-4 double mutants (Severson et al., 2009; Tzur et al., 2012; Severson and Meyer, 2014), demonstrating that both REC-8 and COH-3/4 complexes contribute to sister chromatid cohesion.
Cohesin loading to meiotic chromosomes is not only essential for sister chromatid cohesion, but also for successful completion of key meiotic prophase events. REC-8 and COH-3/4 cohesin complexes associate with axial elements in the transition zone of the germ line, where chromosomes become dramatically elongated compared to mitotic cells (Pasierbek et al., 2001; Hayashi et al., 2007; Nabeshima et al., 2011; Severson and Meyer, 2014). Assembly of axial elements is fully dependent on cohesin loading, as depletion of SMC-1 or SCC-3 (which are common to all meiotic cohesin complexes), lack of all three meiotic kleisins, or lack of the cohesin loading factor SCC-2, prevent association of HORMA-domain proteins with axial elements and cause severe meiotic defects that prevent CO formation (Pasierbek et al., 2003; Wang et al., 2003; Goodyer et al., 2008; Severson et al., 2009; Lightfoot et al., 2011). Loading of different meiosis-specific cohesin complexes is also required for the assembly of axial elements in mice (reviewed in McNicoll et al., 2013). Furthermore, meiotic defects, including impaired chiasma formation, are present in mutants that display a partial reduction in the overall amount of cohesin associated with axial elements, such as in worms carrying a hypomorphic smc-3 allele or mutants lacking the axis-associated LAB-1 protein (Baudrimont et al., 2011; Tzur et al., 2012). CO formation is impaired in mutants lacking either REC-8 or COH-3/4, demonstrating that both types of cohesin play important roles during meiotic recombination (Pasierbek et al., 2001; Hayashi et al., 2007; Severson et al., 2009). Moreover, REC-8 and COH-3/4 cohesin also play different roles in ensuring SC assembly between homologous chromosomes, since SC assembly is greatly reduced in coh-3 coh-4 double mutants and SC installation is thought to occur between sister chromatids in rec-8 mutants (Severson et al., 2009; Severson and Meyer, 2014).
Proteins containing a HORMA domain, a protein motif also found in spindle-assembly checkpoint protein MAD2 (Aravind and Koonin, 1998), play fundamental roles in establishing meiotic chromosome structure. The C. elegans genome encodes four of these proteins—HIM-3, HTP-1, HTP-2, and HTP-3, with unique and redundant functions. Collectively, these HORMA-domain proteins regulate almost all key events of meiosis, ranging from homolog recognition to the correct release of sister chromatid cohesion during the meiotic divisions.
HIM-3 and HTP-1/2 consist mostly of a HORMA domain flanked by short N- and C-terminal regions, while HTP-3 contains a long C-terminal tail that contains four docking sites for HIM-3 and two for HTP-1/2 (Kim et al., 2014). Therefore, HTP-3 is essential for the assembly of axial elements. In addition, HTP-1/2 can also be recruited to axial elements by binding directly to the C-terminal tail of HIM-3 (Kim et al., 2014). HTP-3 is also required to promote loading of REC-8-containing cohesin complexes (Severson et al., 2009) and to mediate the formation of DSBs (Goodyer et al., 2008), both of which are independent of its requirement to recruit HIM-3 and HTP-1/2 (Kim et al., 2014). Interestingly, HTP-3 is the only meiotic HORMA-domain protein that localizes to chromatin in the mitotic compartment of the germ line (Hayashi et al., 2007; Goodyer et al., 2008). Finally, HTP-3 is required for the acquisition of posttranslational modifications on axis-associated chromatin (Couteau and Zetka, 2011). Thus, HTP-3 is not only essential to promote axial element assembly, but may also orchestrate changes in chromatin structure for multiple meiotic prophase events.
HIM-3 and HTP-1 are both required to ensure homolog recognition and to promote faithful SC assembly; as a consequence, him-3 and htp-1 null mutants display severe impairment in CO formation (Zetka et al., 1999; Couteau and Zetka, 2005; Martinez-Perez and Villeneuve, 2005). Although the mechanisms by which these two proteins promote homolog pairing are not well understood, HTP-1 has been proposed to participate in two checkpoint-like mechanisms: the first makes initiation of SC assembly contingent on successful homology recognition, while the second prolongs the homology search process until homolog interactions are stabilized by SC loading (Couteau and Zetka, 2005; Martinez-Perez and Villeneuve, 2005; Silva et al., 2014). A mutant form of HTP-1 that fails to associate with axial elements still supports the delayed exit from early prophase triggered by synapsis defects, suggesting that nuclear soluble HTP-1 may participate in the quality control of SC assembly (Silva et al., 2014). An important difference between htp-1 and him-3 null mutants is that SC assembly is dramatically reduced in him-3 mutants, whereas lack of HTP-1 results in high levels of SC assembly between non-homologous chromosomes. Non-homologous synapsis is also observed in mutants expressing hypomorphic alleles of him-3 (Couteau et al., 2004), demonstrating that both HTP-1 and HIM-3 have regulatory roles in SC assembly. Removal of HTP-2 (which shares 82% identity at the amino acid level with HTP-1) from htp-1 mutants results in a dramatic reduction in SC assembly, demonstrating that HTP-2 can promote homology-independent SC assembly (Couteau and Zetka, 2005; Martinez-Perez et al., 2008). However, the analysis of htp-2 single mutants failed to identify obvious meiotic defects; thus, HTP-2 function appears largely redundant with that of HTP-1 (Martinez-Perez et al., 2008).
In addition to promotion of pairing and synapsis between homologs, HTP-1 and HIM-3 also directly affect meiotic recombination, presumably by making the sister chromatid inaccessible for repair, thereby encouraging repair off the homolog (Couteau et al., 2004; Martinez-Perez and Villeneuve, 2005) (see also Section 5.2.4 and Section 5.5). htp-1 mutants show a reduction in RAD-51 foci (see Section 5) which can be explained by a reduction in the overall number of DSBs formed and/or faster DSB turnover (Couteau and Zetka, 2005; Martinez-Perez and Villeneuve, 2005). Finally, HTP-1 and HTP-2 participate in the two-step release of sister chromatid cohesion during the meiotic divisions, where they act to ensure that a subset of REC-8-containing cohesin remains associated with chromosomes during the first meiotic division (Martinez-Perez et al., 2008; Severson et al., 2009). Of note, the connection between HORMA-domain proteins and the regulation of REC-8 complexes goes beyond the release of sister chromatid cohesion in the meiotic divisions, since htp-1 htp-2 double mutants, him-3 mutants, and rec-8 mutants display premature centriole disengagement during meiosis II in spermatogenesis (Schvarzstein et al., 2013).
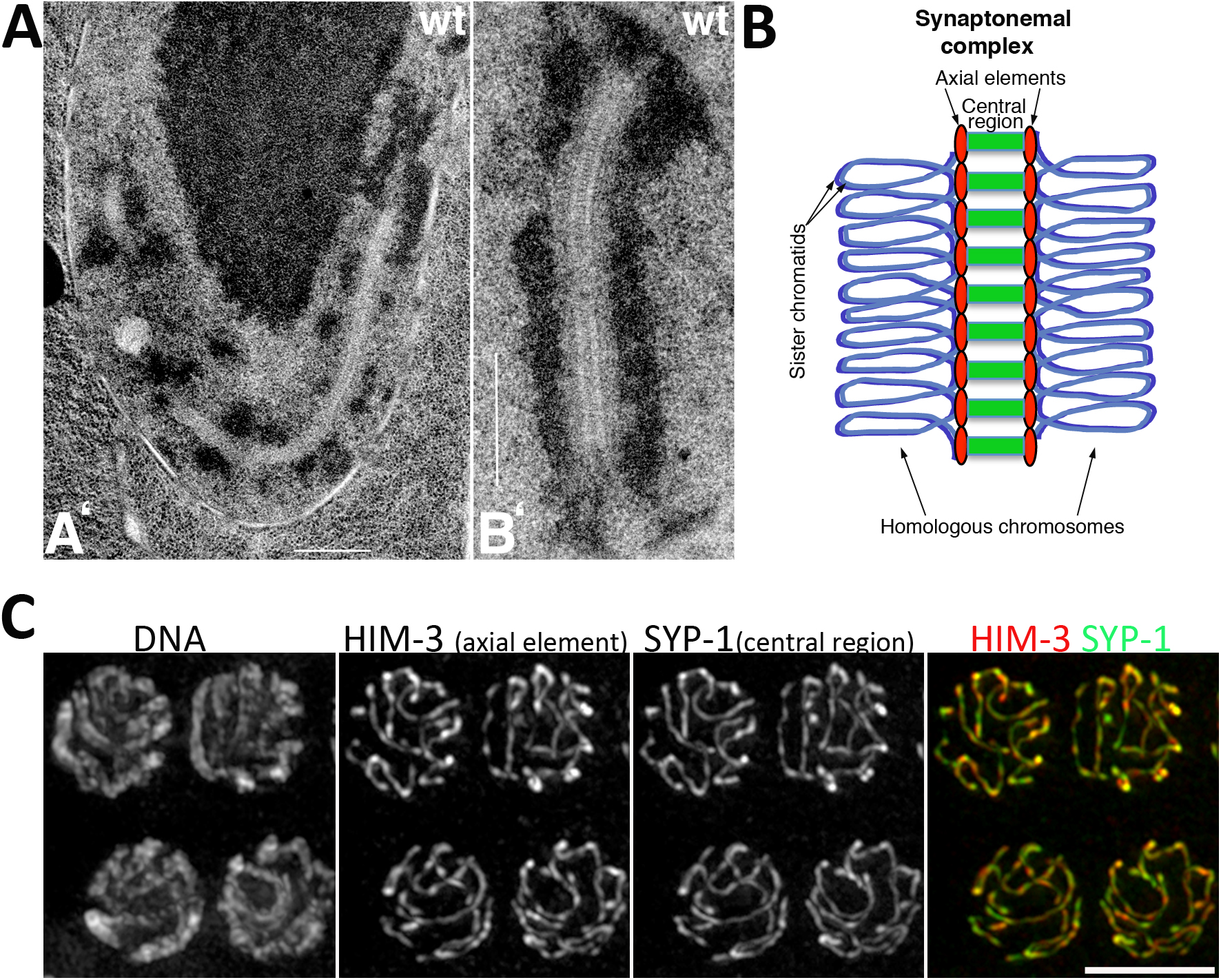
The most prominent, and highly conserved, structural feature of meiotic prophase is the SC. When observed by electron microscopy, the SC appears as a tripartite, ladder-like structure consisting of a central region that contains transverse filaments that bridge the axial elements of paired homologs (MacQueen et al., 2002; Colaiácovo, 2006) (Figure 4). Although structurally similar, with long coiled-coil domains, the major components of the SC in different organisms are poorly conserved at the primary amino acid level. In C. elegans, four components of the central region of the SC have been identified: SYP-1, SYP-2, SYP-3, and SYP-4. Analysis of the corresponding single mutants reveals that these four proteins are interdependent for their loading onto chromosomes; consequently, SC assembly is fully abrogated in each one of the single mutants (MacQueen et al., 2002; Colaiácovo et al., 2003; Smolikov et al., 2007b; Smolikov et al., 2009). SC assembly is also abrogated in htp-3 mutants and in mutants lacking all three meiotic kleisins (REC-8, COH-3, and COH-4) (Goodyer et al., 2008; Severson et al., 2009). Investigation of the interactions among the four SYP proteins by immunoprecipitation, yeast two-hybrid experiments, and immuno-electron microscopy, suggests that SYP-1 and SYP-2 occupy a central position within the SC, while SYP-3 interacts directly with axis components, and SYP-4, the largest of these proteins, interacts with SYP-3 and reaches into the center of the SC (Schild-Prüfert et al., 2011).
|
Figure 4. Structure of the SC. (A). TEM images of nuclei from the late pachytene regions of a wild-type germ line (reproduced with permission from Colaiácovo et al., 2003). (A’) Equatorial section of a wild-type pachytene nucleus containing a very long continuous stretch of SC, visible as a zipper-like track flanked by electron-dense patches of chromatin; the large dark body centered in the upper half of the panel is the nucleolus. (B’) Detail of SC structure from a different wild-type nucleus. Scale bars equal 500 nm. (B) Diagram of SC structure assembled between a pair of homologous chromosomes. Note that each homolog (depicted as a pair of closely associated sister chromatids) assembles an axial element (red), and that central region components (green) link together homologous axial elements. (C) Projections of pachytene nuclei stained with anti-HIM-3 and anti-SYP-1 antibodies, counterstained with DAPI and imaged with a DeltaVision system. Note that axial elements from the homologs cannot be resolved as two separate structures since the distance between them (<100 nm) is below the resolution of conventional fluorescence microscopy (200 nm). Scale bar equals 5 µm.
Once assembled, the SC stabilizes the pairwise association of homologous chromosomes and promotes the formation of interhomolog COs (see also Section 5.2.4 and Section 5.5) (MacQueen et al., 2002; Colaiácovo et al., 2003; Smolikov et al., 2007a; Smolikov et al., 2009). In addition, the SC is required for normal levels of CO interference (see Section 5.4 and Section 5.5) (Hayashi et al., 2010; Libuda et al., 2013) and to promote normal meiotic progression, as evidenced by the persistence of chromosome clustering in syp mutants (see Section 7.1) (MacQueen et al., 2002; Colaiácovo et al., 2003).
SC assembly appears to be a cooperative process and occurs preferentially in a pairwise fashion, linking the axis of two chromosomes (MacQueen et al., 2005; Hayashi et al., 2010; Mlynarczyk-Evans et al., 2013). In vivo imaging of GFP-tagged SYP-3 suggests that synapsis of each homolog pair is initiated only once and that PCs are the primary sites for initiation; once initiated, SC extension continues at a constant rate until full synapsis is achieved (Rog and Dernburg, 2015). Whereas the SC normally assembles only between correctly paired homologs, the SC structure itself is indifferent to homology, as seen by the presence of normal-appearing SC structures between non-homologous chromosomes in some meiotic mutants and in worms heterozygous for chromosome rearrangements (Couteau et al., 2004; Couteau and Zetka, 2005; Martinez-Perez and Villeneuve, 2005; Penkner et al., 2007b; Henzel et al., 2011). Thus, SC assembly must be tightly regulated to ensure a productive outcome. Regulation of SYP protein loading involves axial element components HIM-3 and HTP-1 (see Section 4.2), but proteins that do not become incorporated into the SC structure have also been reported to control the loading of SYP proteins, both in a chromosome-specific and a nucleus-wide manner. For example, the PC-binding HIM-8/ZIM proteins are required for proper assembly of SC in a chromosome-specific manner (Phillips et al., 2005; Phillips and Dernburg, 2006; Rog and Dernburg, 2013) (see also Section 3.2), and in the absence of all PC-binding proteins, SC assembly becomes misregulated and non-homologous SC assembly takes place (Harper et al., 2011; Labella et al., 2011). In addition, the chromosome motions characteristic of early prophase have been shown to be important for proper SC assembly: in their absence, SC assembly often occurs between nonhomologous chromosomes (see Section 3.3). Factors that affect the loading of SYP proteins in a nucleus-wide fashion include: the CHK-2 kinase, required to induce normal levels of SC assembly in early prophase (MacQueen and Villeneuve, 2001; Martinez-Perez and Villeneuve, 2005); HAL-2, a nucleoplasmic protein that prevents the inappropriate loading of SYP proteins, which can interfere with the pairing process (Zhang et al., 2012); LAB-1, a component of axial elements that targets protein phosphatase 1 and is required for full SC assembly (Tzur et al., 2012); CRA-1, the C. elegans homolog of the non-catalytic subunit of the N-terminal acetyltransferases B, which is required to promote normal loading of SYP proteins (Smolikov et al., 2008; Gao et al., 2015); the CSN/COP9 signalosome, which is required to prevent the formation of extrachromosomal aggregates of SC proteins during early prophase (Brockway et al., 2014); and the AAA+- ATPase PCH-2, which localizes to the SC but is dispensable for SC formation and is proposed to prevent synapsis defects by acting as a “brake” that limits the rate of SC assembly to allow time for error correction (Deshong et al., 2014).
SC disassembly starts in late pachytene in a process that is linked to the progression of meiotic recombination (see Section 6). Interestingly, the SC can be locally disassembled in response to exogenous DNA damage (Couteau and Zetka, 2011) or can be completely disassembled from an X chromosome without a CO (Meneely et al., 2012), demonstrating that the SC is a dynamic structure.
In most organisms, including C. elegans, successful chromosome segregation during meiosis requires formation of CO recombination events between homologous chromosomes; these COs, in conjunction with sister chromatid cohesion, serve to tether homologs together. These connections between homologs, in turn, allow correct (bipolar) attachment of homologs to the meiosis I spindle. Release of a subset of cohesin complexes at anaphase I then allows separation of homologs, thereby accomplishing the reduction in ploidy necessary for sexual reproduction.
Meiotic recombination is initiated through introduction of DSBs by the enzyme SPO-11 (Section 5.1). Repair of these DSBs can result in formation of COs (Section 5.2) and the role of the meiotic machinery is to ensure that each chromosome receives at least one CO (the obligate CO). COs do not occur randomly along the chromosomes: each chromosome has a characteristic distribution of COs (Section 5.3). Further, the number of COs per chromosome pair, and their positions relative to each other, are not random, reflecting several types of regulation (Section 5.4). In addition, chromosome structure affects meiotic recombination (Section 5.5), as does the meiotic cell cycle (Section 5.6). See Figure 5 for an overview of the key steps of meiotic recombination.
|
Figure 5. Model depicting the key events of meiotic recombination. Homologous chromosomes are represented in red and blue, and the two sister chromatids of each homolog are represented as pairs of double stranded DNA molecules indicated by two parallel lines in close proximity. Proteins required for specific steps of meiotic recombination are indicated on the left and right hand side of the diagram. Note that the existence of some of the recombination intermediates represented in this diagram has not been directly demonstrated in C. elegans, but are inferred from studies in yeast. The molecular events of recombination are represented in temporal progression starting at the top of the diagram with the formation of a DSB by SPO-11 in a single chromatid of one of the homologs. Resection of DNA ends and RAD-51 loading promote the invasion of a chromatid from the homologous chromosome, the formation of a D loop, and the start of DNA synthesis. These intermediates can be destabilized by the activity of RTEL-1, which leads to repair as NCO products, or they can be stabilized by CO promoting factors that promote the formation of double Holliday junctions. Note that although MSH-4, MSH-5, ZHP-3, and COSA-1 are all required for CO formation and eventually become associated with CO-fated recombination events, there are clear differences in the timing of loading of these proteins that are not depicted on this model (see Section 4.2.3 for a detailed description). The asymmetric cleavage of double Holliday junctions by different endonucleases promotes the formation of inter-homolog CO events.
Recombination initiates with a DSB, introduced by the conserved topoisomerase-like enzyme SPO-11 (Dernburg et al., 1998). Mutants defective in genes required for DSB formation, such as spo-11, manifest 12 univalents at diakinesis because the achiasmate homologs condense and separate from one another during diplotene. A hallmark of this class of mutants is that bivalent formation can be restored by exposing the mutant to sources of exogenous breaks such as ionizing radiation (IR). In addition to spo-11 itself, genes that fall into this category include him-17 (Reddy and Villeneuve, 2004), dsb-1 (Stamper et al., 2013), and dsb-2 (Rosu et al., 2013). Two other genes, xnd-1 (Wagner et al., 2010) and him-5 (Meneely et al., 2012), influence DSB formation mainly on the X chromosome. MRE-11 (Chin and Villeneuve, 2001), RAD-50 (Hayashi et al., 2007), and HTP-3 (Goodyer et al., 2008) are also required for efficient DSB induction, but play additional roles in DSB repair (see below). Finally, CRA-1 regulates histone acetylation and promotes efficient DSB formation (Gao et al., 2015), suggesting that regulation of chromatin structure by histone modifications is an important modulator of DSB, as observed in other organisms.
Repair of DSBs during meiosis shares many features in common with homologous recombination (HR) in mitotic cell cycles, with several meiosis-specific modifications. Following DSB formation, 5’ ending DNA strands are resected, leaving 3’ single-stranded DNA flanking the DSB site. A 3’ end then invades an intact double-stranded template DNA molecule (in meiosis, typically the homologous chromosome rather than the sister chromatid, the preferred repair partner in other contexts) and primes DNA synthesis. Following repair synthesis, this intermediate can be disassembled (e.g., through the action of DNA helicases (Pâques and Haber, 1999)); the released ssDNA end can then re-anneal to the other DSB end, leading to repair of the DSB without CO formation. Alternatively, the intermediate can be stabilized through the action of meiosis-specific CO-promoting factors, leading to formation of more complex, branched CO recombination intermediates (Figure 5). These intermediates must be enzymatically resolved by DNA strand cleavage; this can result in crossing over (reciprocal exchange of information between chromosomes). We depict these meiotic CO-intermediates as double Holliday junctions by analogy to S. cerevisiae, where recombination intermediate structures have been analyzed using physical assays; in most cases, the existence of specific DSB repair intermediates in C. elegans is inferred rather than directly demonstrated.
Mutants defective in genes required for DSB repair cannot be rescued by IR. Instead, exposure of repair-deficient mutants to IR leads to significant chromosomal abnormalities including DNA fragmentation, chromosome aggregation and defects in chromosome condensation, which are readily visible in diakinesis-stage oocytes (e.g., Chin and Villeneuve, 2001).
The first step in the repair of meiotic DSBs is resection, the processing of ends into 3’ single-stranded DNA (ssDNA) overhangs. Based on the high degree of conservation between SPO-11 homologs, it is believed that SPO-11 becomes covalently attached to the DNA upon DSB formation, and that further processing requires removal of the SPO-11-DNA nucleoprotein complexes to create short 3’ overhangs. The MRE-11 5’-3’ exonuclease is then normally required to generate long 3’ ssDNA tails (Yin and Smolikove, 2013) that ultimately get coated with the RAD-51 recombinase (Alpi et al., 2003; Colaiácovo et al., 2003; Petalcorin et al., 2006). In the absence of mre-11, the exonuclease EXO-1 has also been shown to contribute to resection of meiotic DSBs, although resection is then delayed until mid-late pachytene (Yin and Smolikove, 2013).
Mutants defective in resection, such as mre-11 (Chin and Villeneuve, 2001; Yin and Smolikove, 2013) and rad-50 (Hayashi et al., 2007), are characterized by impaired loading of RAD-51 to SPO-11-dependent DSBs and display diakinesis-stage oocytes with abnormal chromosome aggregations. These aggregates arise from repair by non-homologous end joining (NHEJ). The predominance of HR over NHEJ suggests that active mechanisms suppress NHEJ in the meiotic germ line (Smolikov et al., 2007a; Adamo et al., 2010; Lemmens et al., 2013). A key player in this inhibition is COM-1 (the Sae2/CtIP homolog (Penkner et al., 2007a)) that actively inhibits the binding of Ku proteins to the DNA ends (Lemmens et al., 2013). The involvement of MRE-11 in both DSB formation and subsequent resection is another potential mechanism to ensure that DSBs are shunted towards HR.
The RAD-51 nucleoprotein filament serves the important role of promoting the homology search and strand invasion into the homologous duplex DNA (Figure 5). Displacement of the homologous template strand forms a D loop structure wherein the invading 3’ end is used to prime DNA synthesis. COs can result from subsequent formation and cleavage of more complex intermediates (possibly double Holliday junctions) made by second end capture. Non-crossovers (NCOs) are presumed to arise from ejection of the invading strand and repair via synthesis-dependent strand annealing. Failure to execute these steps can allow for aberrant repair via NHEJ and ultimately can cause chromosome fusions.
Due to the central role of RAD-51 to HR, it is no surprise that RAD-51 filament formation is highly regulated (Rinaldo et al., 2002; Alpi et al., 2003; Colaiácovo et al., 2003; Takanami et al., 2003). The worm homolog of the BRCA2 breast cancer associated protein, BRC-2, facilitates nuclear localization of RAD-51, its loading onto ssDNA (Martin et al., 2005), and D loop formation (Petalcorin et al., 2007). In brc-2 mutants, RPA-coated ssDNA can be detected; this either reflects the fact that RPA-1 loads onto resected ssDNA before RAD-51, as in other organisms, or is an aberrant outcome resulting from the failure to load RAD-51. RTEL-1 may also influence RAD-51 dynamics through its role as an anti-recombinase, disrupting D loops and promoting alternative pathways of repair (Barber et al., 2008).
In order for HR to proceed, RAD-51 must also be removed from the D loop. Three proteins have been thought to promote RAD-51 turnover: RAD-54 (Mets and Meyer, 2009), the helicase HELQ-1/HEL308, and the RAD-51 paralog, RFS-1 (Ward et al., 2010), with the latter two having a redundant function in wild-type cells. Cytological examination of both rad-54 single mutants and helq-1 rfs-1 double mutants shows prolonged persistence of RAD-51 filaments, indicating the persistence of unrepaired DSBs.
Once strand exchange has occurred, additional regulation is required to ensure that a subset of these intermediates become committed to a CO fate. Classically, two CO pathways were described in model systems: the class I pathway, requiring the meiosis-specific homologs of the mismatch repair protein MutS, MSH4 and MSH5, and certain SC associated proteins, is considered responsible for COs subject to interference (see Section 5.4); the class II pathway, dependent on the structure-specific nuclease components MUS81 and MMS4, accounts for the interference-independent COs (reviewed in Kohl and Sekelsky, 2013). In C. elegans, the class I pathway predominates, as mutations in the respective orthologs, msh-4, msh-5 (Winand et al., 1998; Zalevsky et al., 1999; Kelly et al., 2000; Colaiácovo et al., 2003), and zhp-3 (Zip3 homologous protein; Jantsch et al., 2004) leads to univalents at diakinesis. It should be noted that MSH-4 and MSH-5 are first detected broadly in presumptive DSB repair foci that exceed the number of eventual COs; they then become restricted to CO sites by late pachytene (Yokoo et al., 2012). ZHP-3 is initially present along the entire SC in mid-pachytene and then gradually becomes restricted to CO sites (Bhalla et al., 2008). Thus, additional proteins must function in designating a subset of recombination intermediates for CO fates. Anti-recombinases such as RTEL function at this step to shunt repair intermediates into a NCO outcome (possibly by promoting synthesis-dependent strand annealing); thus, in the absence of rtel-1 function, additional COs are made (Youds et al., 2010). COSA-1, a cyclin-like protein that is required for CO formation, has been proposed to function in reinforcement of CO designation, as it is predominantly detected as a single focus on each chromosome pair during late pachytene (Yokoo et al., 2012). Additional COSA-1 foci are observed on very large fusion chromosomes, such at the X;IV fusion chromosome mnT12 (Libuda et al., 2013), where multiple COs can be detected genetically (Hillers and Villeneuve, 2003).
Loss of function mutations in MSH-4, MSH-5, ZHP-3, or COSA-1 result in 12 univalents at diakinesis. Irradiation cannot restore bivalent formation in these mutants. The mutants also do not show chromosome fusions indicative of mutations in strand exchange proteins. The kinetics of RAD-51 foci turnover is slower in these mutants, but they do disappear prior to diplotene indicating that DSB repair has occurred (Zalevsky et al., 1999; Colaiácovo et al., 2003; Jantsch et al., 2004; Song et al., 2010; Yokoo et al., 2012).
The emerging picture of CO formation from studies in C. elegans and other systems is that an excess of DSBs are made; these get initially bound by repair proteins and putative CO designation proteins and winnowed down to a small number of CO sites. Understanding this selection process is an area of active research in many organisms including C. elegans.
Although many events of DSB repair are conserved between mitosis and meiosis, a major difference is that the homolog is used as a repair template during meiosis, whereas the sister chromatid is the preferred repair template for cells in S and G2 of mitotic cell cycles. The preference for the homolog rather than the sister is thought to be mediated in large part by axial element components (Section 4.2). Use of the homolog as a recombination partner also appears to be a temporally regulated feature of the meiotic program: access to the homolog as repair template for both CO and non-CO repair is shut down upon transition into the late pachytene stage. Thus, DSBs that are made or persist into late pachytene or beyond are presumed to be repaired using the sister chromatid as a repair template (Colaiácovo et al., 2003; Rosu et al., 2011).
During meiosis, an excess of DSBs is formed (relative to the number of COs). Many of these breaks are repaired as NCOs using the homolog as a repair template (Robert et al., 2008; Rosu et al., 2011), but repair using the sister chromatid as a template likely also occurs. The SMC-5/6 complex and the BRC-1 protein have been shown to be important for repair of meiotic DSBs under conditions where inter-homolog CO formation is abrogated and/or in mutant situations when inter-sister repair is the only option (Boulton et al., 2004; Adamo et al., 2008; Bickel et al., 2010). These papers provided evidence that SMC-5/6 and BRC-1 are important for inter-sister repair, but the data are consistent with these proteins also participating in inter-homolog non-CO repair as well.
The final step of CO recombination is cleavage of CO intermediates to yield mature CO products. C. elegans contains several resolvase functions that are scaffolded by the SLX-4/HIM-18 protein (Saito et al., 2009). These can either act redundantly or can substitute for each other to resolve CO intermediates. XPF-1 (in combination with HIM-6/BLM) and SLX-1 (in combination with MUS-81) are proposed to define two mechanistically distinct pathways for resolving CO intermediates (Agostinho et al., 2013; O'Neil et al., 2013; Saito et al., 2013). Double mutants lacking both pathways exhibit defects in post-pachytene chromosome reorganization and disassembly of the SC. Unique to these mutants, diakinesis univalents appear closely apposed and connected by chromatin bridges (Agostinho et al., 2013; O'Neil et al., 2013; Saito et al., 2013). These bridges are ultimately resolved in meiosis II, however, suggesting that additional resolvase functions are present in the worm germ line.
The HIM-6/BLM helicase may play a central role in ensuring that CO-designated recombination intermediates are eventually resolved into COs. In the absence of HIM-6 protein, nuclei appear to carry out the early steps of meiotic recombination normally; CO designation also seems normal, as 6 COSA-1 foci can be seen in late pachytene oocytes. However, a subset of these CO-designated intermediates fail to form COs, evidenced by the presence of univalents at diakinesis (Schvarzstein et al., 2014). It may be that HIM-6 acts to ensure the CO outcome by biasing resolution of intermediates (by either XPF-1 or SLX-1/MUS-81) towards CO formation or by protecting CO-designated intermediates from NCO resolution.
CO frequency and distribution can be assayed genetically, using either phenotypic markers or DNA sequence polymorphisms; the latter approach allows a more detailed analysis of crossing over, as a higher density of markers can be used in a single experiment (Davis et al., 2005; Hillers and Villeneuve, 2009). Next-generation DNA sequencing has the potential to allow genome-wide determination of the recombination landscape but to date has only been used to probe targeted genomic regions (Kaur and Rockman, 2014). Crossing over can also be analyzed cytologically, by visualizing the number and distribution of COSA-1 foci in late pachytene cells (Yokoo et al., 2012; Libuda et al., 2013).
In many eukaryotes, COs are non-randomly distributed along chromosomes, with a tendency to occur within small regions called hotspots (Nicolas et al., 1989; Baudat et al., 2010). No evidence for CO hotspots has been found in C. elegans, however, despite considerable efforts (Kaur and Rockman, 2014). Nevertheless, meiotic COs in C. elegans are not randomly distributed along chromosomes. As described in the WormBook chapter Karyotype, ploidy and gene dosage, the distribution of COs along the autosomes is uneven, with roughly the central third of each autosome having a paucity of events; interestingly, the same region of each autosome has a higher density of genes (the “central gene cluster”). Conversely, the arms of each chromosome have a higher frequency of crossing over and a lower gene density, thus the physical and genetic maps of C. elegans are said to be “out of alignment”. This type of CO density domain structure is also observed on the X chromosome, but is much less pronounced (Barnes et al., 1995; Rockman and Kruglyak, 2009). Together, these features indicate that some aspect of CO formation is subject to regional regulation; the DNA sequence determinants of this regulation (if any) are unclear.
Evidence that CO distribution is influenced by genetic factors came initially from the isolation of a mutation in the rec-1 gene. This allele alters the distribution of COs along autosomes without changing overall frequency. This redistribution of COs towards the middle of each chromosome aligns the physical and genetic maps of C. elegans (Rose and Baillie, 1979a; Zetka and Rose, 1995). Mutations in the him-5 gene also cause redistribution of meiotic COs along autosomes, resulting in an increased incidence of COs in the central gene cluster; similar to rec-1, there is not an overall increase of COs along autosomes (Meneely et al., 2012). In contrast to rec-1, however, HIM-5 also plays a role in facilitating meiotic DSB formation on the X chromosomes: in a him-5 mutant the frequency of COs on the X is reduced, with a concomitant increase in X chromosome nondisjunction (the Him phenotype).
The phenotypes of him-17, xnd-1, and condensin mutants suggest that chromatin structure and condensation state are also critical determinants of DSB sites. him-17 and xnd-1 mutants both show aberrant histone post-translational modifications and are impaired in DSB formation (Reddy and Villeneuve, 2004; Wagner et al., 2010). Both genes are also required for normal localization of HIM-5, revealing interplay between chromatin and the recombination landscape (Meneely et al., 2012). Likewise, condensin proteins, which play crucial roles in chromosome condensation during cell divisions, play roles in establishing the wild-type distribution of DSBs and COs. Heterozygosity for a null mutation in any of the subunits of Condensin I (DPY-28, DPY-26, MIX-1, SMC-4, CAPG-1) or of KLE-2 (a subunit of Condensin II) leads to HIM-3-dependent alterations in recombination and chromosome structure: COs are redistributed along the X chromosome, the overall frequency of COs is increased, and pachytene chromosome axis length is increased (Tsai et al., 2008; Mets and Meyer, 2009). Mutants homozygous for a partial loss of function allele of dpy-28, which does not increase CO frequencies, showed that CO distribution correlated with a shift in the distribution of RAD-51 foci (Mets and Meyer, 2009). Moreover, the dpy-28 partial loss of function allele suppresses the defect in chiasma formation of him-17 mutants, suggesting that higher-order chromosome structures promoted by condensin limit DSB formation (Tsai et al., 2008). Together, this set of genes points to a complex interplay between chromosome/ chromatin structure and the meiotic recombination machinery.
Despite these general features, CO frequency and position are not fixed within the population, and the number and frequency of COs differs between the sexes, with maternal age, and with temperature (Hodgkin et al., 1979; Rose and Baillie, 1979b; Zetka and Rose, 1990; Lim et al., 2008; Song et al., 2010). However, in all these situations, the arm/cluster differences in CO distribution remain (Barnes et al., 1995). Thus, at the global level, there are defined regions of the genome that are more or less prone to receive a CO.
CO distribution can also be influenced by events subsequent to DSB formation. DSB sites can be mapped cytologically using anti-RAD-51 antibodies in a mutant such as rad-54 that prevents further processing of the filament and effectively traps the recombination intermediate (Mets and Meyer, 2009; Nottke et al., 2011; Rosu et al., 2011; Saito et al., 2012). Cytological examination of RAD-51-marked DSB sites in wild type failed to detect an arm/cluster bias in DSB distribution (Saito et al., 2012), which would be expected if CO distribution were predominantly enforced at the step of DSB formation. Rather, it appears that CO formation in the central gene cluster is actively inhibited at a step post-recruitment of RAD-51. The resolvase SLX-1 (Section 5.2.5) appears to function in this inhibition, as mutation of SLX-1 leads to a redistribution of exchanges along chromosome V: there is an increase in COs in the central gene cluster of chromosome V, with a compensatory decrease in COs in the arm regions (Saito et al., 2012).
Each of the 6 chromosome pairs in C. elegans has a genetic map size near 50 cM. A map size of 50 cM indicates that each chromosome pair experiences (on average) only one CO per meiosis. Each homolog pair must receive the “obligate CO”, to promote accurate chromosome segregation, and different mechanisms operate during meiosis to ensure this outcome. The ability to ensure the formation of at least one CO per homolog pair is known as CO assurance, while the ability to maintain CO number in the face of variability in numbers of DSBs is known as CO homeostasis. As homeostasis is a mechanism that promotes COs at the expense of NCOs in situations where DSB numbers are limiting, CO homeostasis helps to achieve CO assurance, suggesting that these two manifestations of CO regulation are mechanistically linked. Evidence for the operation of these mechanisms during worm meiosis comes from studies in which a limited number of DSBs are exogenously introduced in spo-11 mutants. For example, when a single DSB is generated by transposon excision, this event is converted into a CO with high efficiency (Rosu et al., 2011). This suggests that CO assurance can be achieved by ensuring that each chromosome receives at least one DSB. Similarly, when a limited number of DSBs are generated by irradiation, these are also efficiently converted into a single CO per chromosome pair. Further increases in the number of DSBs by increased irradiation does not result in the formation of more COSA-1 marked events, providing evidence for CO homeostasis (Yokoo et al., 2012). In addition, CO homeostasis may explain why only the X chromosome is devoid of a CO in him-5 mutants whereas RAD-51 foci are reduced over four-fold genome-wide (Meneely et al., 2012).
In C. elegans, most chromosome pairs undergo only a single CO per meiosis (Hillers and Villeneuve, 2003; Hammarlund et al., 2005), but when multiple COs have been mapped, they tend to be widely spaced along the chromosome (Hillers and Villeneuve, 2003; Lim et al., 2008; Libuda et al., 2013). This non-random distribution of COs is known as CO interference (Muller, 1916). The fact that most chromosome pairs receive only a single CO suggests that CO interference operates over distances greater than the length of a typical chromosome to discourage additional exchanges once one has occurred. Indeed, studies with fusion chromosomes have demonstrated that end-to-end fusions of two and even three chromosomes still frequently only received a single CO in meiosis (Hillers and Villeneuve, 2003). Further, cytological mapping of COs along the length of mnT12 (an end-to-end fusion of chromosomes IV and X) enabled measurement of the distance along chromosomes over which CO interference operates in C. elegans, and strengthened the idea that interference operates over distances longer than the length of a typical chromosome (Libuda et al., 2013).
While cytological evaluation of CO interference using COSA-1 foci suggests that chromosome pairs rarely have more that one cytologically differentiated CO site, double COs have been characterized using genetic recombination assays. Double CO data for individual autosomes range between 2-10% per chromosome pair (Meneely et al., 2002; Nabeshima et al., 2004; Lim et al., 2008; Deshong et al., 2014; Gabdank and Fire, 2014). Extrapolating these data for all 5 autosomes, this genetic evidence suggests up to 35% of nuclei have at least one autosome that obtains a second CO. These extra COs are cytologically distinct, however, as they are apparently not associated with COSA-1 foci (Yokoo et al., 2012; Deshong et al., 2014). The X chromosome, in contrast, appears particularly immune to double COs in some studies (Lim et al., 2008; Tsai et al., 2008; but see Deshong et al., 2014), perhaps due to its heterochromatic-like state (Kelly et al., 2002). Further, the frequency of double COs differs between oocyte and sperm meiosis (Zetka and Rose, 1990; Lim et al., 2008; Gabdank and Fire, 2014). Data are only available for male sperm: they appear to have less stringent genetic interference compared to oocytes, having both increased numbers of double COs and more closely spaced COs (Lim et al., 2008; Gabdank and Fire, 2014).
While the analysis of null mutants lacking any of the SYP proteins reveals that these proteins are required for CO/chiasma formation, partial depletion of SYP proteins by RNAi results in an increased incidence of double COs (Hayashi et al., 2010; Libuda et al., 2013). Similarly, a non-null allele of him-3 that allows partial synapsis of homologs also results in increased incidence of double COs (Nabeshima et al., 2004). This apparent contradiction can be explained by the proposal that the SC functions in the process of CO interference (Section 5.4). Although the mechanism by which interference is transmitted along chromosomes is not understood, Libuda et al. (2013) found that each CO event is associated with a local extension of the axial element, suggesting that this structural alteration may play a role in impeding the formation of further COs. CO numbers are also increased when the structure of chromosomes is altered by reducing the dose of components of the condensin I complex, although in this case the extra COs are not associated with a COSA-1 marked cytologically differentiated site (Tsai et al., 2008; Mets and Meyer, 2009; Yokoo et al., 2012).
DSB formation and repair are initiated in the transition zone; RAD-51 foci continue to accumulate through mid-pachytene, suggesting that both the initiation and cessation of DSB formation are controlled. DSB formation has been proposed to be subject to feedback regulation; once all chromosome pairs have a CO-eligible intermediate, DSB formation ceases (Section 7.2). In addition, it is clear that as nuclei transition into late pachytene, there is a shift in repair mechanism utilization; even if DSBs are formed in late pachytene germ cells (e.g., by gamma irradiation), these nuclei are incapable of committing to CO formation (Yokoo et al., 2012). Further, during early and mid-pachytene, RAD-50 is required for RAD-51 loading at DSB sites; after the late pachytene transition, RAD-50 is no longer required for RAD-51 loading (Hayashi et al., 2007). Finally, access to the homolog as a repair template is also shut down at the transition from mid- to late pachytene (Rosu et al., 2011), presumably reflecting reversion to preferential use of the sister chromatid as a repair partner. This transition to sister repair is accompanied by an ability of DNA damage to trigger localized separation of the chromosome axes, a feature that is antagonized by HTP-3 at earlier stages (Couteau and Zetka, 2011). Ultimately, this transition in DNA repair mode may allow for residual DSBs to be repaired and the nucleus to escape apoptosis induced by the DNA damage checkpoint. The transition zone to mid-pachytene region can thus be considered the window of opportunity for the formation of DSBs and CO-designated events.
Following the formation of CO-fated recombination intermediates, meiotic chromosomes undergo two major structural changes in preparation for the meiotic divisions: the disassembly of the SC, which occurs in an asymmetric fashion around the CO site; and condensation, a process that causes a dramatic reduction in chromosome size (for detailed reviews see Schvarzstein et al., 2010; Wood et al., 2010). Together, these lead to the formation of diakinesis bivalents, highly condensed pairs of homologous chromosomes held together by sister chromatid cohesion and COs. Bivalents contain two functional domains: a short arm where sister chromatid cohesion will be released during the first meiotic division, and a long arm where sister chromatids will remain attached until the onset of the second meiotic division (Albertson and Thomson, 1993; Albertson et al., 1997).
The creation of bivalents with two differentiated functional domains starts in late pachytene with the redistribution of SC and axis-associated proteins that is triggered by CO-fated recombination intermediates (Nabeshima et al., 2005; de Carvalho et al., 2008; Martinez-Perez et al., 2008; Agostinho et al., 2013) (Figure 6). At this stage HTP-1/2 and LAB-1 are depleted from the region of the chromosomes between the CO site and the closest telomere, forming the short arm of the bivalent, while the SYP proteins and ZHP-3 are depleted from the region between the CO and the farthest telomere, forming the long arm (Nabeshima et al., 2005; Smolikov et al., 2007b; Bhalla et al., 2008; de Carvalho et al., 2008; Martinez-Perez et al., 2008). COH-3/4-containing cohesin complexes are also largely removed from the long arm of each bivalent by late diakinesis and become restricted to the short arm by prometaphase I (Severson and Meyer, 2014). The short arm region undergoes a further change in protein composition in the two most proximal oocytes in which SYP proteins are removed and AIR-2 kinase is recruited (Kaitna et al., 2002; Rogers et al., 2002; Nabeshima et al., 2005). AIR-2 recruitment to diakinesis bivalents is required for homolog segregation during the first meiotic division (Kaitna et al., 2002; Rogers et al., 2002). REC-8 is phosphorylated by AIR-2 in vitro (Rogers et al., 2002), suggesting that REC-8 phosphorylation may be important for cohesion release in vivo. In fact, REC-8 staining in the short arm of bivalents becomes weaker during late diakinesis and prometaphase (Rogers et al., 2002; de Carvalho et al., 2008; Harper et al., 2011; Severson and Meyer, 2014), and this reduction in REC-8 staining requires AIR-2 (Severson and Meyer, 2014). By metaphase I, REC-8 staining is greatly reduced in the short arm of bivalents (de Carvalho et al., 2008), but use of a different anti-REC-8 antibody shows clearly detectable REC-8 staining in the short arm of most metaphase I bivalents (Pasierbek et al., 2001; Rogers et al., 2002;; Cortes et al., 2015). REC-8 and COH-3/4 cohesin complexes that remain bound on the short arm of metaphase I chromosomes must be removed at anaphase onset. A likely candidate for this activity is the conserved protease separase, which is required for proper meiotic chromosome segregation (Siomos et al., 2001). Interestingly, separase is not required for the late prophase removal of REC-8 and COH-3/4 cohesin described above (Severson and Meyer, 2014), suggesting that distinct mechanisms mediate cohesin removal during C. elegans meiosis. Phosphorylation of kleisin subunits is required for their cleavage by separase in yeast mitotic and meiotic cells (reviewed in Haarhuis et al., 2014), and it has been proposed that AIR-2-dependent phosphorylation of REC-8 complexes may render them sensitive to separase (Rogers et al., 2002; de Carvalho et al., 2008). AIR-2 activity in the long arms of diakinesis bivalents is antagonized by GSP-2 (protein phosphatase 1), whose depletion leads to the premature separation of sister chromatids during anaphase I (Kaitna et al., 2002; Rogers et al., 2002). Similarly, HTP-1/2 and LAB-1 prevent the inappropriate recruitment of AIR-2 to the long arm of the bivalents, and their absence causes untimely removal of REC-8 and premature release of sister chromatid cohesion during the first meiotic division (de Carvalho et al., 2008; Martinez-Perez et al., 2008). Since the ability of LAB-1 to antagonize AIR-2 localization depends on the presence of GSP-2, and since LAB-1 and GSP-2 physically interact, LAB-1 likely acts by recruiting GSP-2 to chromosomes (de Carvalho et al., 2008; Tzur et al., 2012).
|
Figure 6. Asymmetric re-distribution of SC components during bivalent differentiation. Top row shows projections of individual bivalents taken from nuclei at the indicated stages and stained with anti-HTP-1/2 and anti-SYP-1 antibodies and counterstained with DAPI. The bottom row shows a diagram of the process of bivalent differentiation. During pachytene HTP-1/2 and SYP-1 colocalize along the whole length of fully synapsed bivalents, but by late pachytene HTP-1/2 and SYP-1 acquire a reciprocal staining pattern, with a CO (or a CO-fated event) marking the boundary of the two domains. SC disassembly and chromosome condensation start during diplotene, when the regions lacking SYP proteins become separated. By diakinesis, bivalents undergo further condensation and the long arms, containing HTP-1/2, and short arms, containing SYP-1, are clearly differentiated. For a detailed description of bivalent differentiation see Section 5.1.
Concomitant with the changes in protein composition described above, the overall structure of chromosomes is modified by a process of chromatin compaction that starts in late pachytene and continues until the onset of the first meiotic division. This process is mediated by condensin complexes, which are structurally similar to cohesin, containing a core of two SMC proteins that associate with additional factors to modify the topology of chromosomes (reviewed in Wood et al., 2010). The condensin II complex (MIX-1, SMC-4, HCP-6, CAPG-2, KLE-2) associates with chromosomes in diplotene and is required for the compaction and resolution of chromosomes during diplotene and diakinesis, demonstrating that condensin II is essential for the remodeling of chromosomes during late meiotic prophase (Hagstrom et al., 2002; Chan et al., 2004; Csankovszki et al., 2009; Mets and Meyer, 2009). In diakinesis oocytes, condensin II decorates chromatin on the long and short arms of the bivalents. On the other hand, depletion of condensin I (MIX-1, SMC-4, DPY-26, DPY-28, CAPG-1) does not induce gross defects in the condensation or individualization of meiotic chromosomes during late prophase, and condensin I components are only observed associated with chromosomes on the short arm of the most proximal oocyte, loading only after nuclear envelope breakdown (Tsai et al., 2008; Csankovszki et al., 2009; Meyer, 2010; Collette et al., 2011; also see X-chromosome dosage compensation). During the meiotic divisions, depletion of condensin I components induces mild defects that are only evident during the second meiotic division (Csankovszki et al., 2009), while depletion of condensin II induces dramatic defects in both meiotic divisions, including chromatin bridges between separating chromosomes in anaphase I and II (Chan et al., 2004; Csankovszki et al., 2009). Thus, the reshaping of chromosomes by condensin during late prophase is an essential aspect of meiosis.
The formation of COs during prophase I of meiosis is essential to promote the accurate segregation of chromosomes into the gametes. Therefore, multiple processes that promote the formation of COs, including the establishment of the SC and the progression of recombination, are closely monitored. Failures of those processes lead to a meiotic progression delay for the correction of errors and/or the induction of apoptosis to eliminate meiocytes where errors persist.
The assembly of the SC plays a central role in mediating meiotic progression. The absence of synapsis on one or more chromosomes causes nuclei to delay exit from zygotene, resulting in a prolongation of chromosome movement and chromosome clustering (MacQueen et al., 2002; Phillips et al., 2005; Carlton et al., 2006; Phillips and Dernburg, 2006; Penkner et al., 2009; Sato et al., 2009; Baudrimont et al., 2010). This delay requires HTP-1: htp-1; syp-2 double mutants have no zygotene arrest, leading to the proposal that HTP-1 participates in the generation of an inhibitory signal that blocks exit from zygotene until the SC is installed between all homologous pairs (Martinez-Perez and Villeneuve, 2005). This inhibitory signal appears to involve a soluble pool of HTP-1, since a mutant version of HTP-1 that fails to associate with axial elements still supports zygotene arrest in SC-deficient mutants (Silva et al., 2014). SC assembly appears to directly antagonize the inhibitory signal that blocks early meiotic progression, as improper SC assembly between sister chromatids in syp-3(me42) mutants results in normal meiotic progression (Smolikov et al., 2007b).
CHK-2 and PLK-2 protein kinases have essential roles in organizing and monitoring features of chromosome end mobilization (Harper et al., 2011; Labella et al., 2011). Coincident with chromosome movements in leptotene/zygotene, SUN-1 shows prominent phosphorylation at several residues in its nuclear amino-terminus that are dependent on CHK-2 and PLK-2 (Penkner et al., 2009; Harper et al., 2011; Labella et al., 2011). SUN-1 serine-12 phosphorylation is restricted to SUN-1 molecules concentrated at PCs, whereas SUN-1 phosphorylated at other residues is found throughout the entire nuclear envelope (Penkner et al., 2009). Although PC mobilization still takes place when SUN-1 is rendered non-phosphorylatable, synapsis proceeds at a slower rate. In addition, SUN-1 phosphorylation is required to sustain PLK-2 recruitment to PC attachments in the absence of synapsis; this suggests that phosphorylation of SUN-1 can prolong the time window of chromosome movement (Woglar et al., 2013). The interdependency of SUN-1 phosphorylation by PLK-2 and PLK-2 recruitment by phospho-SUN-1 therefore establishes a positive feedback loop to sustain chromosome end-led mobility for as long as necessary to complete synapsis.
In addition to prolonging PC-mediated chromosome movement, the presence of unsynapsed PCs also triggers apoptosis during oocyte meiosis. Induction of this apoptosis by unpaired PCs requires PCH-2 (Bhalla and Dernburg, 2005). Synapsis at PCs is monitored with the help of MES-4 and MET-1 (MES-4 alone for the X chromosome) (Lamelza and Bhalla, 2012).
In males (chromosomally XO), the single X chromosome remains partnerless during meiosis and thus needs to be shielded from the mechanisms that recognize asynapsed chromosomes and induce delays in meiotic progression (Jaramillo-Lambert et al., 2010). The single X chromosome in males assumes a condensed state and is enriched for repressive chromatin modifiers that are important for shielding the single X from the checkpoint machinery (Checchi and Engebrecht, 2011). Transient pseudosynapsis of X chromosome sister chromatids has been observed and has been proposed to mask the X from being recognized as partnerless (Checchi et al., 2014). Furthermore, SUN-1 aggregates at the single X chromosome in males are not phosphorylated by PLK-2, despite its localization to the X PC (Woglar et al., 2013).
Successful completion of meiosis requires formation of COs between each chromosome pair; this in turn requires formation of at least one DSB per homolog pair. However, DSBs also represent possible sources of damage to the genome, and thus, C. elegans has multiple mechanisms to monitor formation of DSBs and downstream recombination intermediates in order to limit the numbers of DSBs that form, to assure that sufficient DSBs are generated to ensure CO formation, and to prevent germ cells with persistent DNA damage from proceeding to the oocyte meiotic divisions.
A growing body of evidence from multiple organisms suggests that CO intermediates function as a rheostat for the DSB machinery, maintaining DSB formation when levels are low and turning it off when DSBs are high (Lange et al., 2011; Gray et al., 2013; Kauppi et al., 2013; Thacker et al., 2014; Garcia et al., 2015). In C. elegans, evidence for such a feedback system comes from analysis of SUN-1 and from DSB-1 and DSB-2 (Rosu et al., 2013; Stamper et al., 2013), two proteins that are required for efficient DSB formation and that localize to chromosomes during early meiotic prophase concomitant with DSB formation. Similar to SUN-1 phosphorylation, DSB-1/2 chromatin loading depends on CHK-2 (Rosu et al., 2013; Stamper et al., 2013). In synapsis- or recombination-defective mutants, the zone of SUN-1 phosphorylation and DSB-1/2 localization is extended (Stamper et al., 2013; Woglar et al., 2013), suggesting the presence of a checkpoint-like feedback system that monitors formation of CO intermediates and retains nuclei in a DSB permissive state until enough CO-fated events are formed. Failure to form a CO intermediate between the X chromosomes (as seen in him-5 and him-8 mutants) results in delayed meiotic progression, suggesting that this feedback system is capable of detecting the absence of crossing over (or a CO-fated intermediate) on a single chromosome pair (Meneely et al., 2012; Stamper et al., 2013). (Checchi et al., 2014) found that the ATM-1 protein kinase plays a conserved role in limiting DSB formation, as mutants defective in atm-1 have elevated levels of RAD-51 foci; however, whether ATM-1 activation is part of the above-described feedback mechanism or represents a distinct mechanism for limiting DSB numbers remains an open question.
During oogenesis, cells with unrepaired recombination intermediates are subjected to apoptosis dependent on the DNA damage sensors MRT-2, HPR-9, HUS-1, and CEP-1, the p53 homolog (Gartner et al., 2000; Schumacher et al., 2001); (also see Germline survival and apoptosis). Mutants with high levels of resected single stranded DNA, such as rad-51, fcd-2, brc-1, or brc-2, elicit massive apoptosis (Gartner et al., 2000; Martin et al., 2005; Adamo et al., 2008; Adamo et al., 2010). Interestingly, the pro-CO factors MSH-4, MSH-5, and ZHP-3 are required for the induction of apoptosis irrespective of DNA damage checkpoint activation, as evaluated by the transcriptional activation of egl-1 (Silva et al., 2013). The ability of unrepaired meiotic DSBs to trigger DNA damage-checkpoint-mediated apoptosis also provides a means to cull nuclei that had entered meiotic prophase with aneuploidy accumulated during the preceding mitotic divisions (Stevens et al., 2013).
In males, where apoptosis is not triggered, many features of damage sensing and signaling nevertheless are activated and thereby contribute to faithful processing of recombination intermediates (Jaramillo-Lambert et al., 2010).
After meiotic entry (which is covered in Germline proliferation and its control) oocytes remain in G2 throughout their progression along the gonad. Progression through prophase I must be coordinated with the cell cycle and oocyte growth for proper gamete development. prom-1 mutants enter meiosis and express axis and SC proteins. However, the processes of pairing and synapsis are defective: few nuclei display the asymmetrical chromatin distribution characteristic of the transition zone (leptotene/zygotene), loading of axis and SC proteins is delayed, and the synapsis that eventually does occur is often between nonhomologous chromosomes (Jantsch et al., 2007). One known function of the SCFprom-1 ubiquitin ligase is the degradation of CYE-1, which plays a role in the germline proliferative zone (Fox et al., 2011). It is therefore possible prom-1 connects meiotic events to cell cycle progression.
ERK/MAP kinase MPK-1 is required for progression to late pachytene; cells lacking MPK-1 never enter a diplotene-like state (Church et al., 1995; Lackner and Kim, 1998). This is consistent with accumulation of the phosphorylated—hence activated—form of MPK-1 in mid-pachytene (Lee et al., 2007). The transition from homolog-directed DSB repair to sister-directed repair, with the accompanying loss of RAD-50 dependence for RAD-51 loading (Section 5.6), depends on MPK-1 (Hayashi et al., 2007). A key determinant of the length of prophase during C. elegans oogenesis is the translational repressor LIN-41, which coordinates oocyte growth with meiotic maturation. In lin-41 mutants pachytene cells prematurely cellularize, diplotene does not occur, and oocytes prematurely enter M phase. This premature M phase entry is elicited in part by translational derepression of a CDK-1 inhibitor (Spike et al., 2014). For further reading on oocyte maturation and oocyte release from prophase arrest, see Control of oocyte meiotic maturation and fertilization as well as Kim et al., 2013.
Proper segregation of chromosomes at meiosis I requires orientation of homologs toward opposite spindle poles and the localized release of sister chromatid cohesion from the short arms of the bivalents at anaphase onset (see Section 6.1). During metaphase I, outer kinetochore components are seen forming two opposing cup-like structures that encase each half of the bivalent (Howe et al., 2001; Monen et al., 2005). This organization promotes the mono-orientation of sister chromatids on the spindle, since sister chromatids of the long arm of the bivalent are encased within the same cup-like structure and thus behave as a single unit. Mono-orientation of sister chromatids in metaphase I also requires normal sister chromatid cohesion, as rec-8 mutants show premature separation of sisters at anaphase I (Severson et al., 2009).
During oogenesis, meiotic chromosome segregation takes place in the context of a barrel-shaped acentriolar spindle that needs to be correctly positioned, in close association with the cortex, within the large volume of the fertilized embryo (Albertson and Thomson, 1993). Proper spindle assembly in oocytes requires MEI-1 and MEI-2, the components of the microtubule severing complex Katanin (Mains et al., 1990; Clark-Maguire and Mains, 1994; Srayko et al., 2000), the kinesin KLP-18 (Segbert et al., 2003), and the calponin homology-domain protein ASPM-1 (van der Voet et al., 2009; Connolly et al., 2014). Correct positioning of the meiotic spindle in oocytes is a two-step process that involves translocation of the spindle to the cortex, which occurs before metaphase I, followed by a 90° rotation that renders the spindle perpendicular to the cortex before the onset of anaphase I (reviewed in Fabritius et al., 2011). Spindle translocation requires kinesin-1, MEI-1, and microtubules (Yang et al., 2003; Yang et al., 2005), while spindle rotation is promoted by the anaphase promoting complex, dynein, and a complex formed by LIN-5, ASPM-1, and CMD-1 (Yang et al., 2005; Ellefson and McNally, 2009; van der Voet et al., 2009).
Detailed imaging of microtubules and kinetochores during meiosis I shows that bivalents are surrounded by robust microtubule bundles running along their sides, and that the mid-bivalent region is surrounded by a ring-like structure containing the chromosome passenger complex and the chromokinesin KLP-19 (Wignall and Villeneuve, 2009). This suggests that lateral associations with microtubules (rather than canonical end-on kinetochore attachments) facilitate chromosome movements prior to anaphase. Although kinetochores are required for proper chromosome orientation during congression, potentially helping to align chromosomes within the lateral bundles, the migration of chromosomes during anaphase I is kinetochore independent (Dumont et al., 2010). Instead, lateral microtubule associations remain intact during anaphase to facilitate segregation (Muscat, et.al. 2015). At anaphase onset, the mid-bivalent ring structure containing KLP-19 is removed from chromosomes and is left behind in the center of the spindle (Dumont et al., 2010), allowing minus-end forces to drive poleward movement along lateral bundles (Muscat, et.al. 2015). Moreover, spindle elongation also facilitates separation, potentially stimulated by another ring component, CLS-2/CLASP (Dumont et al., 2010). Kinetochores are disassembled shortly after the onset of anaphase I, and are reassembled by metaphase II. In meiosis II, each sister chromatid is encased in a cup-like kinetochore, and KLP-19 and the chromosome passenger complex localize to a ring-like structure that encircles the region of remaining contact between the two sister chromatids (Wignall and Villeneuve, 2009; Dumont et al., 2010).
The processes that mediate meiotic chromosome segregation differ between oogenesis and spermatogenesis (Albertson and Thomson, 1993; Shakes et al., 2009; Wignall and Villeneuve, 2009; Schvarzstein et al., 2013). In contrast to oocyte meiotic spindles, spermatocyte meiotic spindles assume a more traditional configuration. They have centrioles at their poles and the metaphase spindles have high microtubule density adjacent to kinetochore domains at the ends of the bivalents, indicative of end-on attachments between microtubules and kinetochores. Furthermore, anaphase separation of chromosomes in spermatocytes is driven by separation of spindle poles. Finally, completion of the meiotic divisions produces markedly different outcomes in spermatogenesis, where four functional haploid gametes are produced, and oogenesis, where one haploid gamete and two degenerate polar bodies are formed (also see Control of oocyte meiotic maturation and fertilization). Interestingly, the asymmetry of oocyte meiosis can partially correct for pre-existing trisomies, since in triplo-X animals the extra X chromosome is preferentially extruded into the first polar body, thus reducing the incidence of aneuploid progeny (Cortes et al., 2015).
C. elegans has proven to be a very productive organism for the study of meiosis. The excellent cytology, coupled with the ease of isolation of meiotic mutants, has contributed to our understanding of the complex series of events necessary to produce haploid gametes. However, understanding the molecular mechanisms that regulate homolog pairing, meiotic recombination, the changes in structure that chromosomes undergo during meiotic prophase, the process of chromosome segregation, and the quality control of meiosis remain important goals for future studies. The recent development of efficient transgenic tools, combined with super-resolution microscopy, long-term in vivo imaging of meiotic chromosomes, rapid methods to map recombination events, and the use of proteomic approaches means that C. elegans will remain an excellent model organism to address these questions over the coming years.
We thank Barbara Meyer, Anne Villeneuve, and David Greenstein for critically reading the manuscript and members of the Jantsch, Martinez-Perez, and Yanowitz labs for comments and discussions. We are grateful to Patricia Hamminger, Sophia Millonigg, Alexander Woglar, and Sarah Testori for figure preparation. The authors labs were funded by the FWF (VJ), an MRC core funded grant (E M-P), NIH (JLY/VJ & KJH), and the March of Dimes (JLY).
Genes are organized in alphabetical order and numbers in brackets indicate the section(s) within the main text where the functions of a given protein are discussed.
| Gene Name | Ortholog/Homolog | Molecular Function (known or inferred) | Subcellular Localization (if known) | Meiotic Role |
|---|---|---|---|---|
| air-2 | Aurora/ IPL Kinase | Kinase | Recruited to the short arm of diakinesis bivalents | Loss impairs chromosome segregation at meiosis I; phosphorylates REC-8 in vitro. (Section 6.1) |
| aspm-1 | ASPM | Interaction with calmodulin and microtubules | Meiotic spindles | Required for proper spindle assembly in oocytes and to promote spindle rotation in a complex with CMD-1 and LIN-5. (Section 8) |
| brc-1 | BRCA1 | DNA repair | Required for intersister DSB repair (Section 5.2.4); mutation results in high levels of resected ssDNA and massive germline apoptosis (Section 7.2). | |
| brc-2 | BRCA2 | DNA repair | Facilitates nuclear entry of RAD-51, loading of RAD-51 on ssDNA, and D loop formation (Section 5.2.2). Mutation results in high levels of resected ssDNA and massive germline apoptosis (Section 7.2). | |
| capg-1 | hCAP-G | Condensin I complex | Nuclear localization in mitotic and meiotic prophase nuclei. Associates with short arm of bivalents during late diakinesis and metaphase I, and between sister chromatids at metaphase II | Heterozygous null mutants show altered distribution of DSBs and COs, increased CO frequencies, and increased pachytene axis length (Section 5.3). Required for chromosome segregation (Section 6.2). |
| capg-2 | hCAP-G2 | Condensin II complex | Decorates chromatin in diakinesis oocytes and during metaphase I and II | Required for chromosome condensation and segregation. (Section 6.2) |
| cep-1 | p53 | DNA binding, transciptional activation | DNA damage sensing, involved in DNA damage-induced apoptosis in the germ line. (Section 7.2) | |
| chk-2 | CHK2 | Kinase | Master regulator of early prophase events. Required for loading of autosomal PC proteins (ZIMs); required for formation of SUN-1 aggregates (Section 3.2). Required for homolog pairing and normal loading of SC in early prophase (Section 4.3.1). Required for N-terminal phosphorylation of SUN-1 shortly after meiotic entry (Section 7.1). Required for DSB formation and loading of DSB-1/2 (Section 7.2). | |
| coh-3 | Rad21L/Rec8 | Meiosis-specific kleisin subunit of cohesin complex | Chromosome axes | COH-3 and COH-4 are highly identical and functionally redundant. Loss of COH-3 and COH-4 impairs CO formation and SC assembly. Loss of COH-3, COH-4, and REC-8 causes gross defects in sister chromatid cohesion and prevents assembly of axial elements. (Section 4.1) |
| coh-4 | Same as coh-3 | Same as coh-3 (above). | Same as coh-3 (above). | Same as coh-3 (above). |
| com-1 | CtlP/Sae2 | DNA repair; resection of DSBs | Promotes processing of SPO-11 DSBs to facilitate RAD-51 loading; suppresses NHEJ. (Section 5.2.1) | |
| cosa-1 | CTND1 | CO designation | Late pachytene—one focus per chromosome pair (presumed to mark CO position) | Designates recombination intermediates for CO formation; required for CO formation. (Section 5.2.3) |
| cra-1 | NAA25 | N(alpha)-acetyl-transferase | Localizes to meiotic nuclei throughout prophase | Required to promote normal loading of SC proteins (Section 4.3.1) and for regulation of histone acetylation and DSB formation (Section 5.1). |
| cye-1 | CCNE1 | E-type cyclin | Promotes mitotic proliferation in the distal germ line. Negatively regulated in meiotic cells by PROM-1 ubiquitin ligase. (Section 7.3) | |
| dpy-26 | hCAP-H | Condensin I complex | Nuclear localization in mitotic and meiotic prophase nuclei. Associates with short arm of metaphase I bivalents and between sister chromatids at metaphase II | Heterozygous null mutants show altered distribution of DSBs and COs, increased CO frequencies, and increased pachytene axis length (Section 5.3). Required for chromosome segregation (Section 6.2). |
| dpy-28 | hCAP-D2 | Condensin I complex | Same as dpy-26 (above). | Heterozygotes same as dpy-26 (above). Homozygous mutants of hypomorph allele show changes in distribution of DSBs and COs without altering CO frequency (Section 5.3). Required for chromosome segregation (Section 6.2). |
| dsb-1 | Localizes to chromatin in transition zone and early pachytene nuclei | Involved in DSB formation; regulates competence for DSB formation (Section 5.1). Localizes to chromosomes, dependent on CHK-2. Localization prolonged in synapsis or recombination mutants; suggests a role in maintenance of DSB-permissive state (Section 7.2). | ||
| dsb-2 | Same as dsb-1 (above). | Same as dsb-1 (above). Loss of DSB-2 from chromosomes may correlate with switch from homolog to sister repair (Section 7.2). | ||
| egl-1 | Positive regulation of apoptosis | Positive regulator of apoptosis. (Section 7.2) | ||
| exo-1 | exo1 | DNA repair; exonuclease | Involved in resection of meiotic DSBs. (Section 5.2.1) | |
| fcd-2 | FANCD2 | DNA repair | Mutations result in high levels of resected ssDNA and massive germline apoptosis. (Section 7.2) | |
| gsp-2 | PP1CA | Protein Phosphatase I | Antagonizes AIR-2 recruitment to chromosomes and physically interacts with LAB-1; loss leads to premature sister chromatid separation at Anaphase I. (Section 6.1) | |
| hal-2 | Localized to the nucleoplasm in all germline nuclei. | Promotes homolog pairing, prevents inappropriate loading of SYP proteins; required for DSB formation; required for mechanism that couples cessation of chromosome movement with SC assembly. (Section 4.3.1) | ||
| hcp-6 | hCAP-D3 | Condensin II complex | Associates with chromatin from diplotene, persisting during metaphase I and II | Required for chromosome condensation and segregation. (Section 6.2) |
| helq-1 | HELQ | DNA repair; helicase | Promotes RAD-51 turnover (removal of RAD-51 from D loops); redundant with RFS-1. (Section 5.2.2) | |
| him-3 | HORMAD1; Hop1 | HORMA-domain protein; Component of chromosome axis | Localizes to axial elements throughout meiotic prophase | HORMA domain flanked by short N- and C- termini; required for homolog recognition. Promotes use of homolog as DSB repair template; absence leads to dramatic reduction in SC. Required to prevent premature centriole disengagement in spermatocyte meiosis II (Section 4.2.) Non-null mutations can affect chromosome axis integrity and regulation of meiotic recombination (Section 5.5). |
| him-5 | promotion of DSB formation | Localizes to autosomes from mitotic region through late pachytene | Required for normal DSB formation on X chromosomes. mutation reduces DSB levels 4-fold, but only X typically lacks CO. Required for normal distribution of COs genome-wide (Section 5.1, Section 5.3, Section 5.4). In absence, chromosome clustering/mobility extended and meiotic progression delayed (Section 7.2). | |
| him-6 | BLM; RecQ | DNA repair; Holliday junction resolvase | Resolvase activity (in conjunction with XPF-1). Thought to enforce crossover-biased resolution of CO-designated intermediates (Section 5.2.5) | |
| him-8 | X chromosome pairing and synapsis | Binds X pairing center | Required for homolog alignment and synapsis of X. Recruitment site for polo-like kinases PLK-2/PLK-1 (Section 3.2, Section 4.3). | |
| him-17 | Promotion of DSB formation | Localizes to chromatin throughout the germ line | Mutants accumulate aberrant histone modifications and display impaired DSB formation. Required for HIM-5 localization to chromosomes. (Section 5.1, Section 5.3) | |
| him-18 | SLX4 | DNA repair; Holliday junction resolution | Scaffold for resolvases. (Section 5.2.5) | |
| him-19 | Involved in homolog recognition, defects in him-19 mutants worsen with age. (Section 3.3) | |||
| hpr-9 | RAD9 | DNA damage sensing; 9-1-1 complex | DNA damage sensing, involved in DNA damage-induced apoptosis in the germ line. (Section 7.2) | |
| htp-1 | HORMAD1; Hop1 (yeast) | HORMA-domain protein; Component of chromosome axis | Localizes to axial elements throughout meiotic prophase, but becomes restricted to the long arms of bivalents during late pachytene. Large pool also present as nuclear soluble protein. | Same as HTP-2 (below), except HTP-1 is required for homolog recognition, to prevent non-homologous synapsis, and for CO formation. Promotes use of homolog as DSB repair template (Section 4.2). Thought to help generate inhibitory signal that blocks zygotene exit until all homologous pairs have SC installed (Section 7.1). |
| htp-2 | HORMAD1; Hop1 (yeast) | HORMA-domain protein; Component of chromosome axis | Localizes to axial elements throughout meiotic prophase, but becomes restricted to the long arms of bivalents during late pachytene. | HORMA domain flanked by short N- and C- termini. HTP-1 and HTP-2 are 82% identical; HTP-2 is largely redundant with HTP-1. Together HTP-1 and HTP-2 are required for regulated cohesion release during meiotic divisions (Section 6.1) and to prevent premature centriole disengagement during meiosis II in spermatogenesis (Section 4.2). Loss of HTP-2 does not affect pairing or CO formation (Section 4.2). |
| htp-3 | HORMAD1; Hop1 (yeast) | HORMA-domain protein; Component of chromosome axis | Localizes to nuclei in the mitotic compartment of the germ line. Localizes to axial elements throughout meiotic prophase | HORMA domain flanked by short N-terminus and long C-terminus. Component of meiotic chromosome axis, required for efficient DSB induction. Required for loading of HIM-3, HTP-1/2, and REC-8 cohesin, for SC assembly, and for acquisition of posttranslational modifications on axis-associated chromatin. (Section 4.2, Section 4.3). Promotes DSB formation (Section 5.1). Antagonizes separation of axial elements in early-mid pachytene (Section 5.6). |
| hus-1 | HUS1 | DNA damage sensing; 9-1-1 complex | Localizes to chromatin following DNA damage | DNA damage sensing, involved in DNA damage-induced apoptosis in the germ line. (Section 7.2) |
| kle-2 | hCAP-H2 | Condensin II complex | Decorates chromatin in diakinesis oocytes and during metaphase I and II | Heterozygous null mutants show altered distribution of DSBs and COs, increased CO frequencies, and increased pachytene axis length (Section 5.3). Required for chromosome condensation and segregation. (Section 6.2) |
| klp-18 | Kinesin | Meiotic spindles; higher concentration at spindle poles | Required for assembly of acentrosomal spindles in oocytes. (Section 8) | |
| klp-19 | Chromokinesin | Forms ring-like structure around mid-bivalent region in meiosis I, and around region of remaining contact between sister chromatids in meiosis I | Mediates anti-poleward chromosome movement via lateral microtubules. (Section 8) | |
| lab-1 | Targets GSP-2 to chromosomes | Associates with axial elements and becomes restricted to the long arms of diakinesis bivalents | Axial element component. Targets phosphatase 1 (GSP-2) to chromosomes and antagonizes AIR-2 recruitment (Section 6.1). Required for full SC assembly and for normal sister chromatid cohesion (Section 4.1, Section 4.3.1). Regulates release of sister chromatid cohesion during the first meiotic division (Section 6.1). | |
| lin-41 | Translational repressor | Present, mostly in cytoplasm, from mid pachytene until oocyte maturation | Coordinates oocyte growth with meiotic maturation. Required to prevent premature entrance into M phase. (Section 7.3) | |
| mei-1 | KATNAL1 | Microtubule severing | Spindles and chromosomes during the meiotic divisions | With MEI-2 forms the katanin microtubule severing complex. Required for proper assembly and positioning of meiotic spindle in oocytes. (Section 8) |
| mei-2 | Microtubule severing | Localizes in the cytoplasm of late diakinesis oocytes and to spindles and chromatin during meiotic divisions | Same as for mei-1 (above). | |
| mes-4 | WHSC1; SEDB1 | Histone methyltransferase | Localizes to autosomes in mitotic region, late pachytene, and oocytes | Involved in monitoring synapsis of autosomes and X chromosomes. (Section 7.1) |
| mix-1 | SMC2 | Condensin I and II complexes | Associates with chromatin from diplotene (condensin II), and with the short arm of metaphase I bivalents and between sister chomatids during metaphase II (condensin I) | Heterozygous null mutants show altered distribution of DSBs and COs, increased CO frequencies, and increased pachytene axis length (Section 5.3). Required for chromosome condensation and segregation (Section 6.2). |
| met-1 | SETD2 | Histone methyltransferase | Involved in monitoring synapsis of autosomes. (Section 7.1) | |
| met-2 | SETDB1 | Histone methyltransferase | Shields univalent X in males from synapsis checkpoint. (Section 7.1) | |
| mpk-1 | ERK | MAP kinase | Phosphorylated form (active) present in mid pachytene | Required for progression to late pachytene; mutants never enter diplotene-like state. Regulates transition in mode of DSB repair. (Section 7.3) |
| mre-11 | MRE11A | DNA repair. Nuclease; forms complex with RAD-50 | Required for efficient formation of meiotic DSBs and for RAD-51 loading to SPO-11 DSBs (Section 5.1, Section 5.2.1). | |
| mrg-1 | Chromo-domain protein | Localizes to autosomes | Plays a role in non-pairing center mediated homolog alignment. (Section 3.2) | |
| msh-4 (him-14) | MSH4 | DNA repair; CO designation | Early pachytene - multiple foci per chromosome. Late pachytene, one focus per chromosome (presumed to mark CO position) | Forms a heterodimer with MSH-5. Stabilizes CO intermediates to promote CO outcomes (Section 5.2.3). Required for induction of apoptosis (Section 7.2). |
| msh-5 | MSH5 | DNA repair; CO designation | Same as msh-4 (above). | Same as msh-4 (above). |
| mus-81 | MUS81 | DNA repair; Holliday junction resolvase | Resolvase activity (in conjunction with SLX-1). (Section 5.2.5) | |
| pch-2 | TRIP13 | AAA-ATPase | Forms foci in germline nuclei before transition zone, and SC-like tracks during pachytene | Restricts SC assembly: SC assembly accelerated in mutants (Section 4.3.1). Required for induction of apoptosis when synapsis fails (Section 7.1). |
| plk-1 | PLK1 | Polo-like kinase | Localizes to pairing centers, specially in absence of PLK-2 | Can partially replace meiotic functions of PLK-2 in promoting pairing and SC assembly. (Section 3.2) |
| plk-2 | PLK1 | Polo-like kinase | Localizes to pairing centers in early pachytene and to the SC in mid pachytene | Induces nuclear envelope reorganization; SUN-1 and ZYG-12 relocate into aggregates at clustered PC ends of chromosomes. Required for N-terminal phosphorylation of SUN-1 shortly after meiotic entry (Section 3.2). Involved in feedback loop to sustain chromosome end-led mobility until completion of synapsis (Section 7.1). |
| prom-1 | FBXO47 | F-box protein | Required for homolog pairing, normal assembly of SC, DSB repair; involved in degradation of CYE-1. (Section 7.3) | |
| mrt-2 | RAD1 | DNA damage sensing; 9-1-1 complex | DNA damage sensing, involved in DNA damage-induced apoptosis in the germ line. (Section 7.2) | |
| rad-50 | RAD50 | DNA repair; forms complex with MRE-11 | Required for formation and repair of meiotic DSBs (Section 5.1, Section 5.2.1). Required for RAD-51 loading to DSBs (and interhomolog CO formation) (Section 5.2.1), but only in early to mid-pachytene; in late pachytene, RAD-50 is not required for repair (which is preferentially targeted to sister) (Section 5.6, Section 7.3). | |
| rad-51 | Rad51; DMC1 | DNA repair; catalyzes strand invasion | Localizes to sites of DSBs | Promotes DNA strand invasion (Section 5.2.1, Section 5.2.2, Section 5.6). Mutation results in high levels of resected ssDNA and massive germline apoptosis (Section 7.2). |
| rad-54 | Rad54 | DNA repair; DEAD-like helicase family | Promotes removal of RAD-51 from recombination intermediates (Section 5.2.2). Mutation traps DSBR intermediates as RAD-51 filaments (Section 5.3). | |
| rec-1 | Required for normal distribution of COs (but not frequency) (Section 5.3). | |||
| rec-8 | Rec8 | Meiosis specific kleisin subunit of cohesin complex | Chromosome axes | Required for CO formation and to prevent SC formation between sister chromatids (Section 4.1). Loss of REC-8, COH-3, and COH-4 impairs axial element assembly and causes gross defects in sister chromatid cohesion (Section 4.1). Regulated REC-8 removal during late prophase and metaphase I required for two step-release of cohesion (Section 6.1). Required for mono-orientation of sister chromatids on the first meiotic spindle (Section 8). Required to prevent premature centriole disengagement during meiosis II in spermatogenesis (Section 4.2). |
| rfs-1 | RAD51C | DNA repair; interacts with RAD-51 | RAD-51 paralog; Promotes RAD-51 turnover (removal of RAD-51); redundant with helq-1 (Section 5.2.2). | |
| rpa-1 | RPA1 | DNA repair; single-strand binding protein | Binds to ssDNA at DSB sites in brc-2 mutants; may also bind in WT. (Section 5.2.2) | |
| rtel-1 | RTEL1 | DNA repair; DNA helicase | Anti-recombinase: disrupts D loops in vitro, promotes alternative forms of repair (most likely SDSA; shunts a subset of DSBR intermediates into NCO repair) (Section 5.2.2, Section 5.3). | |
| scc-2 | SCC2 | Cohesin loader | Mutants fail to load to cohesin and to assemble axial elements and the SC. (Section 4.1) | |
| scc-3 | SCC3 | Cohesin component | Chromosome axes | Common to all cohesin complexes. Required for loading of axial element and SC components. (Section 4.1) |
| slx-1 | Resolvase activity (in conjunction with MUS-81) (Section 5.2.5). Inhibits CO formation in the central gene clusters of autosomes (Section 5.3). | |||
| smc-1 | Chromosome axes | Same as scc-3 (above). | ||
| smc-3 | Chromosome axes | Common to all cohesin complexes. Hypomorph allele causes reduction in axis-associated cohesin (Section 4.1). | ||
| smc-4 | SMC4 | Condensin I and II complexes | Associates with chromatin from diplotene (condensin II), and with the short arm of metaphase I bivalents and between sister chomatids during metaphase II (condensin I) | Heterozygous null mutants show altered distribution of DSBs and COs, increased CO frequencies, and increased pachytene axis length (Section 5.3). Required for chromosome condensation and segregation (Section 6.2). |
| smc-5 | SMC5 | Forms complex with SMC-6 | Involved in intersister DSB repair and/or interhomolog NCO repair. (Section 5.2.4) | |
| smc-6 | SMC6 | Forms complex with SMC-5 | Same as smc-6 (above). | |
| spd-3 | Mitochondrial | Required for normal chromosome movements during meiosis; mutation results in non-homologous synapsis. (Section 3.3) | ||
| spo-11 | SPO11 | Formation of meiotic DSBs | Catalyzes formation of meiotic double-strand breaks (Section 3.2, Section 5.1, Section 5.4). | |
| sun-1 | SUN-domain family | Transmembrane protein, interaction with KASH-domain proteins | Nuclear envelope, forms aggregates in transition zone nuclei | Binds lamins and KASH domain proteins forming aggregates in transition zone nuclei at sites of chromosome attachment. Required for homolog pairing and recombination (Section 3.2). Shows CHK-2 and PLK-2 - dependent phosphorylation at N terminus during leptotene-zygotene. Phospho-SUN-1 required for normal rate of SC assembly (slower in absence) and to sustain PLK-2 recruitment to chromosome ends (Section 7.1, Section 7.2). Phospho SUN-1 persists into pachytene in absence of DSBs or CO intermediates (Section 7.2). |
| syp-1 | Synaptonemal complex central region component | Localizes along chromosomes during prophase I | Same as syp-2 (below). Additionally, partial depletion alters CO distribution (Section 5.5). | |
| syp-2 | Synaptonemal complex central region component | Localizes along chromosomes during prophase I | Structural component of synaptonemal complex required for loading of other SYPs and for CO formation. Stabilizes interhomolog interactions during pachytene; promotes interhomolog DSB repair and normal meiotic progression (Section 4.3, Section 7.1). | |
| syp-3 | Synaptonemal complex central region component | Localizes along chromosomes during prophase I | Same as syp-2 (above). Also may interact directly with axis components (Section 4.3). Allele syp-3(me42) leads to SC assembly between sisters (Section 7.1). | |
| syp-4 | Synaptonemal complex central region component | Localizes along chromosomes during prophase I | Same as syp-2 (above). Interacts with SYP-3, spans into center of SC (Section 4.3). | |
| tim-1 | TIMELESS | Cohesin loading | Nuclear localization in mitotic compartment of the germ line and in diplotene and diakinesis oocytes | Required for loading of REC-8 cohesin complexes. (Section 4.1) |
| unc-116 | Kinesin 1 | Kinesin | Required for translocation of the oocyte spindle to the cortex of the embryo. (Section 8) | |
| xnd-1 | AT-hook containing protein | localizes to autosomes from mitotic region through late pachytene | Required for subset of DSBs (mostly on X) (Section 5.1). Mutants accumulate aberrant histone modifications, and result in impaired DSB formation. Required for HIM-5 localization to chromosomes (Section 5.3). | |
| xpf-1 | ERCC4 | DNA repair; Nucleotide excision repair | Resolvase activity (in conjunction with HIM-6). (Section 5.2.5) | |
| zhp-3 | RNF212; Zip3 (yeast) | RING finger protein; possible SUMO ligase | Early pachytene - broadly distributed along SC. End of pachytene, diplotene - one focus per chromosome pair (presumed to mark CO position) | Required for CO formation, role in stabilizing CO-fated intermediates (Section 5.2.3). Also required for induction of apoptosis irrespective of DNA damage checkpoint activation (Section 7.2). |
| zim-1 | Zinc-finger protein | Pairing center of chromosomes II and III in early prophase nuclei | Required for homolog alignment and synapsis of II and III. Binds to PC ends only from leptotene to early pachytene. Localization depends on CHK-2. Recruitment site for polo-like kinases PLK-2/PLK-1. (Section 3.2) | |
| zim-2 | Zinc-finger protein | Pairing center of chromosome V in early prophase nuclei | Same as zim-1 (above) except is required for homolog alignment and synapsis of V. | |
| zim-3 | Zinc-finger protein | Pairing center of chromosomes I and IV in early prophase nuclei | Same as zim-1 (above) except is required for homolog alignment and synapsis of I and IV. | |
| zyg-12 | KASH-domain family | Interacts with SUN-1 in nuclear envelope-spanning protein complex | Nuclear envelope, forms aggregates in transition zone nuclei | Connects nucleus to cytoskeleton (via dynein). Binds SUN-1. Required for homolog pairing and recombination (Section 3.2). Recruits Dynein motor complex to cytoplasmic side of SUN-1/ZYG-12 aggregates in leptotene/zygotene; facilitates proper synapsis (Section 3.3). |
Adamo, A., Collis, S.J., Adelman, C.A., Silva, N., Horejsi, Z., Ward, J.D., Martinez-Perez, E., Boulton, S.J., and La Volpe, A. (2010). Preventing nonhomologous end joining suppresses DNA repair defects of Fanconi anemia. Mol. Cell 39, 25-35. Abstract Article
Adamo, A., Montemauri, P., Silva, N., Ward, J.D., Boulton, S.J., and La Volpe, A. (2008). BRC-1 acts in the inter-sister pathway of meiotic double-strand break repair. EMBO Rep. 9, 287-292. Abstract Article
Agostinho, A., Meier, B., Sonneville, R., Jagut, M., Woglar, A., Blow, J., Jantsch, V., and Gartner, A. (2013). Combinatorial regulation of meiotic Holliday junction resolution in C. elegans by HIM-6 (BLM) helicase, SLX-4, and the SLX-1, MUS-81 and XPF-1 nucleases. PLoS Genet. 9, e1003591. Abstract Article
Albertson, D.G., Rose, A.M., and Villeneuve, A.M. (1997). Chromosome organization, mitosis, and meiosis. In C. Elegans II, D.L. Riddle, T. Blumenthal, B.J. Meyer, and J. Preiss, eds. (Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press), pp. 47-78. Abstract
Albertson, D.G., and Thomson, J.N. (1993). Segregation of holocentric chromosomes at meiosis in the nematode, Caenorhabditis elegans. Chromosome Res. 1, 15–26. Abstract Article
Alpi, A., Pasierbek, P., Gartner, A., and Loidl, J. (2003). Genetic and cytological characterization of the recombination protein RAD-51 in Caenorhabditis elegans. Chromosoma 112, 6–16. Abstract Article
Aravind, L., and Koonin, E.V. (1998). The HORMA domain: a common structural denominator in mitotic checkpoints, chromosome synapsis and DNA repair. Trends Biochem. Sci. 23, 284–286. Abstract Article
Barber, L.J., Youds, J.L., Ward, J.D., McIlwraith, M.J., O'Neil, N.J., Petalcorin, M.I., Martin, J.S., Collis, S.J., Cantor, S.B., Auclair, M., et al. (2008). RTEL1 maintains genomic stability by suppressing homologous recombination. Cell 135, 261–271. Abstract Article
Barnes, T.M., Kohara, Y., Coulson, A., and Hekimi, S. (1995). Meiotic recombination, noncoding DNA and genomic organization in Caenorhabditis elegans. Genetics 141, 159–179. Abstract
Baudat, F., Buard, J., Grey, C., Fledel-Alon, A., Ober, C., Przeworski, M., Coop, G., and de Massey, B. (2010). PRDM9 is a major determinant of meiotic recombination hotspots in humans and mice. Science 327, 836–40. Abstract Article
Baudrimont, A., Penkner, A., Woglar, A., Machacek, T., Wegrostek, C., Gloggnitzer, J., Fridkin, A., Klein, F., Gruenbaum, Y., Pasierbek, P., et al. (2010). Leptotene/zygotene chromosome movement via the SUN/KASH protein bridge in Caenorhabditis elegans. PLoS Genet. 6, e1001219. Abstract Article
Baudrimont, A., Penkner, A., Woglar, A., Mamnun, Y.M., Hulek, M., Struck, C., Schnabel, R., Loidl, J., and Jantsch, V. (2011). A new thermosensitive smc-3 allele reveals involvement of cohesin in homologous recombination in C. elegans. PLoS One 6, e24799. Abstract Article
Bhalla, N., and Dernburg, A.F. (2005). A conserved checkpoint monitors meiotic chromosome synapsis in Caenorhabditis elegans. Science 310, 1683–1686. Abstract Article
Bhalla, N., Wynne, D.J., Jantsch, V., and Dernburg, A.F. (2008). ZHP-3 acts at crossovers to couple meiotic recombination with synaptonemal complex disassembly and bivalent formation in C. elegans. PLoS Genet. 4, e1000235. Abstract Article
Bickel, J.S., Chen, L., Hayward, J., Yeap, S.L., Alkers, A.E., and Chan, R.C. (2010). Structural maintenance of chromosomes (SMC) proteins promote homolog-independent recombination repair in meiosis crucial for germ cell genomic stability. PLoS Genet. 6, e1001028. Abstract Article
Boulton, S.J., Martin, J.S., Polanowska, J., Hill, D.E., Gartner, A., and Vidal, M. (2004). BRCA1/BARD1 orthologs required for DNA repair in Caenorhabditis elegans. Curr. Biol. 14, 33–39. Abstract Article
Brockway, H., Balukoff, N., Dean, M., Alleva, B., and Smolikove, S. (2014). The CSN/COP9 signalosome regulates synaptonemal complex assembly during meiotic prophase I of Caenorhabditis elegans. PLoS Genet. 10, e1004757. Abstract Article
Carlton, P.M., Farruggio, A.P., and Dernburg, A.F. (2006). A link between meiotic prophase progression and crossover control. PLoS Genet. 2, e12. Abstract Article
de Carvalho, C.E., Zaaijer, S., Smolikov, S., Gu, Y., Schumacher, J.M., and Colaiácovo, M.P. (2008). LAB-1 antagonizes the Aurora B kinase in C. elegans. Genes Dev. 22, 2869–2885. Abstract Article
Chan, R.C., Chan, A., Jeon, M., Wu, T.F., Pasqualone, D., Rougvie, A.E., and Meyer, B.J. (2003). Chromosome cohesion is regulated by a clock gene paralogue TIM-1. Nature 423, 1002–1009. Abstract Article
Chan, R.C., Severson, A.F., and Meyer, B.J. (2004). Condensin restructures chromosomes in preparation for meiotic divisions. J. Cell Biol. 167, 613–625. Abstract Article
Checchi, P.M., and Engebrecht, J. (2011). Caenorhabditis elegans histone methyltransferase MET-2 shields the male X chromosome from checkpoint machinery and mediates meiotic sex chromosome inactivation. PLoS Genet. 7, e1002267. Abstract Article
Checchi, P.M., Lawrence, K.S., Van, M. V, Larson, B.J., and Engebrecht, J. (2014). Pseudosynapsis and decreased stringency of meiotic repair pathway choice on the hemizygous sex chromosome of Caenorhabditis elegans males. Genetics 197, 543–560. Abstract Article
Chin, G.M., and Villeneuve, A.M. (2001). C. elegans mre-11 is required for meiotic recombination and DNA repair but is dispensable for the meiotic G(2) DNA damage checkpoint. Genes Dev. 15, 522–534. Abstract Article
Church, D.L., Guan, K.L., and Lambie, E.J. (1995). Three genes of the MAP kinase cascade, mek-2, mpk-1/sur-1 and let-60 ras, are required for meiotic cell cycle progression in Caenorhabditis elegans. Development 121, 2525–2535. Abstract
Clark-Maguire, S., and Mains, P.E. (1994). mei-1, a gene required for meiotic spindle formation in Caenorhabditis elegans, is a member of a family of ATPases. Genetics 136, 533–546. Abstract
Colaiácovo, M.P. (2006). The many facets of SC function during C. elegans meiosis. Chromosoma 115, 195–211. Abstract Article
Colaiácovo, M.P., MacQueen, A.J., Martinez-Perez, E., McDonald, K., Adamo, A., La Volpe, A., and Villeneuve, A.M. (2003). Synaptonemal complex assembly in C. elegans is dispensable for loading strand-exchange proteins but critical for proper completion of recombination. Dev. Cell 5, 463–474. Abstract Article
Collette, K.S., Petty, E.L., Golenberg, N., Bembenek, J.N., and Csankovszki, G. (2011). Different roles for Aurora B in condensin targeting during mitosis and meiosis. J. Cell Sci. 124, 3684–3694. Abstract Article
Connolly, A.A., Osterberg, V., Christensen, S., Price, M., Lu, C., Chicas-Cruz, K., Lockery, S., Mains, P.E., and Bowerman, B. (2014). Caenorhabditis elegans oocyte meiotic spindle pole assembly requires microtubule severing and the calponin homology domain protein ASPM-1. Mol. Biol. Cell 25, 1298–1311. Abstract Article
Cortes, D.B., McNally, K.L., Mains, P.E., and McNally, F.J. (2015). The asymmetry of female meiosis reduces the frequency of inheritance of unpaired chromosomes. eLife 4, e06056. Abstract Article
Couteau, F., Nabeshima, K., Villeneuve, A., and Zetka, M. (2004). A component of C. elegans meiotic chromosome axes at the interface of homolog alignment, synapsis, nuclear reorganization, and recombination. Curr. Biol. 14, 585–592. Abstract Article
Couteau, F., and Zetka, M. (2005). HTP-1 coordinates synaptonemal complex assembly with homolog alignment during meiosis in C. elegans. Genes Dev. 19, 2744–2756. Abstract Article
Couteau, F., and Zetka, M. (2011). DNA damage during meiosis induces chromatin remodeling and synaptonemal complex disassembly. Dev. Cell 20, 353–363. Abstract Article
Csankovszki, G., Collette, K., Spahl, K., Carey, J., Snyder, M., Petty, E., Patel, U., Tabuchi, T., Liu, H., McLeod, I., et al. (2009). Three distinct condensin complexes control C. elegans chromosome dynamics. Curr. Biol. 19, 9–19. Abstract Article
Davis, M.W., Hammarlund, M., Harrach, T., Hullett, P., Olsen, S., and Jorgensen, E.M. (2005). Rapid single nucleotide polymorphism mapping in C. elegans. BMC Genomics 6, 118. Abstract Article
Dernburg, A.F., McDonald, K., Moulder, G., Barstead, R., Dresser, M., and Villeneuve, A.M. (1998). Meiotic recombination in C. elegans initiates by a conserved mechanism and is dispensable for homologous chromosome synapsis. Cell 94, 387–398. Abstract Article
Deshong, A.J., Ye, A.L., Lamelza, P., and Bhalla, N. (2014). A quality control mechanism coordinates meiotic prophase events to promote crossover assurance. PLoS Genet. 10, e1004291. Abstract Article
Dombecki, C.R., Chiang, A.C.Y., Kang, H.-J., Bilgir, C., Stefanski, N.A., Neva, B.J., Klerkx, E.P., and Nabeshima K. (2011). The chromodomain protein MRG-1 facilitates SC-independent homologous pairing during meiosis in Caenorhabditis elegans. Dev. Cell 21, 1092–1103. Abstract Article
Dumont, J., Oegema, K., and Desai, A. (2010). A kinetochore-independent mechanism drives anaphase chromosome separation during acentrosomal meiosis. Nat. Cell Biol. 12, 894–901. Abstract Article
Ellefson, M.L., and McNally, F.J. (2009). Kinesin-1 and cytoplasmic dynein act sequentially to move the meiotic spindle to the oocyte cortex in Caenorhabditis elegans. Mol. Biol. Cell 20, 2722–2730. Abstract Article
Fabritius, A.S., Ellefson, M.L., and McNally, F.J. (2011). Nuclear and spindle positioning during oocyte meiosis. Curr. Opin. Cell Biol. 23, 78–84. Abstract Article
Fox, P.M., Vought, V.E., Hanazawa, M., Lee, M.-H., Maine, E.M., and Schedl, T. (2011). Cyclin E and CDK-2 regulate proliferative cell fate and cell cycle progression in the C. elegans germline. Development 138, 2223–2234. Abstract Article
Fridkin, A., Mills, E., Margalit, A., Neufeld, E., Lee, K.K., Feinstein, N., Cohen, M., Wilson, K.L., and Gruenbaum, Y. (2004). Matefin, a Caenorhabditis elegans germ line-specific SUN-domain nuclear membrane protein, is essential for early embryonic and germ cell development. Proc. Natl. Acad. Sci. U. S. A. 101, 6987–6992. Abstract Article
Gabdank, I., and Fire, A.Z. (2014). Gamete-type dependent crossover interference levels in a defined region of Caenorhabditis elegans chromosome V. G3 4, 117–120. Abstract
Gao, J., Kim, H.-M., Elia, A.E., Elledge, S.J., and Colaiácovo, M.P. (2015). NatB domain-containing CRA-1 antagonizes hydrolase ACER-1 linking acetyl-CoA metabolism to the initiation of recombination during C. elegans meiosis. PLoS Genet. 11, e1005029. Abstract Article
Garcia, V., Gray, S., Allison, R.M., Cooper, T.J., and Neale, M.J. (2015). Tel1(ATM)-mediated interference suppresses clustered meiotic double-strand-break formation. Nature 520, 114–118. Abstract Article
Gartner, A., Milstein, S., Ahmed, S., Hodgkin, J., and Hengartner, M.O. (2000). A conserved checkpoint pathway mediates DNA damage-induced apoptosis and cell cycle arrest in C. elegans. Mol. Cell 5, 435–443. Abstract Article
Goldstein, P. (1982). The synaptonemal complexes of Caenorhabditis elegans: pachytene karyotype analysis of male and hermaphrodite wild-type and him mutants. Chromosoma 86, 577–593. Abstract Article
Goodyer, W., Kaitna, S., Couteau, F., Ward, J.D., Boulton, S.J., and Zetka, M. (2008). HTP-3 links DSB formation with homolog pairing and crossing over during C. elegans meiosis. Dev. Cell 14, 263–274. Abstract Article
Gray, S., Allison, R.M., Garcia, V., Goldman, A.S.H., and Neale, M.J. (2013). Positive regulation of meiotic DNA double-strand break formation by activation of the DNA damage checkpoint kinase Mec1(ATR). Open Biol. 3, 130019. Abstract Article
Haarhuis, J.H.I., Elbatsh, A.M.O., and Rowland, B.D. (2014). Cohesin and its regulation: on the logic of X-shaped chromosomes. Dev. Cell 31, 7–18. Abstract Article
Hagstrom, K.A., Holmes, V.F., Cozzarelli, N.R., and Meyer, B.J. (2002). C. elegans condensin promotes mitotic chromosome architecture, centromere organization, and sister chromatid segregation during mitosis and meiosis. Genes Dev. 16, 729–742. Abstract Article
Hammarlund, M., Davis, M.W., Nguyen, H., Dayton, D., and Jorgensen, E.M. (2005). Heterozygous insertions alter crossover distribution but allow crossover interference in Caenorhabditis elegans. Genetics 171, 1047–1056. Abstract Article
Harper, N.C., Rillo, R., Jover-Gil, S., Assaf, Z.J., Bhalla, N., and Dernburg, A.F. (2011). Pairing centers recruit a Polo-like kinase to orchestrate meiotic chromosome dynamics in C. elegans. Dev. Cell 21, 934–947. Abstract Article
Hayashi, M., Chin, G.M., and Villeneuve, A.M. (2007). C. elegans germ cells switch between distinct modes of double-strand break repair during meiotic prophase progression. PLoS Genet. 3, e191. Abstract Article
Hayashi, M., Mlynarczyk-Evans, S., and Villeneuve, A.M. (2010). The synaptonemal complex shapes the crossover landscape through cooperative assembly, crossover promotion and crossover inhibition during Caenorhabditis elegans meiosis. Genetics 186, 45–58. Abstract Article
Henzel, J. V, Nabeshima, K., Schvarzstein, M., Turner, B.E., Villeneuve, A.M., and Hillers, K.J. (2011). An asymmetric chromosome pair undergoes synaptic adjustment and crossover redistribution during Caenorhabditis elegans meiosis: implications for sex chromosome evolution. Genetics 187, 685–699. Abstract Article
Herman, R.K., and Kari, C.K. (1989). Recombination between small X chromosome duplications and the X chromosome in Caenorhabditis elegans. Genetics 121, 723–737. Abstract
Hillers, K.J., and Villeneuve, A.M. (2003). Chromosome-wide control of meiotic crossing over in C . elegans. Curr. Biol. 13, 1641–1647. Abstract Article
Hillers, K.J., and Villeneuve, A.M. (2009). Analysis of meiotic recombination in Caenorhabditis elegans. Methods Mol. Biol. 557, 77–97. Abstract Article
Hodgkin, J., Horvitz, H.R., and Brenner, S. (1979). Nondisjunction mutants of the nematode Caenorhabditis elegans. Genetics 91, 67–94. Abstract
Howe, M., McDonald, K.L., Albertson, D.G., and Meyer, B.J. (2001). HIM-10 is required for kinetochore structure and function on Caenorhabditis elegans holocentric chromosomes. J. Cell Biol. 153, 1227–1238. Abstract Article
Jantsch, V., Pasierbek, P., Mueller, M.M., Schweizer, D., Jantsch, M., and Loidl, J. (2004). Targeted gene knockout reveals a role in meiotic recombination for ZHP-3, a Zip3-related protein in Caenorhabditis elegans. Mol. Cell. Biol. 24, 7998–8006. Abstract Article
Jantsch, V., Tang, L., Pasierbek, P., Penkner, A., Nayak, S., Baudrimont, A., Schedl, T., Gartner, A., andLoidl, J. (2007). Caenorhabditis elegans prom-1 is required for meiotic prophase progression and homologous chromosome pairing. Mol. Biol. Cell 18, 4911–4920. Abstract Article
Jaramillo-Lambert, A., Ellefson, M., Villeneuve, A.M., and Engebrecht, J. (2007). Differential timing of S phases, X chromosome replication, and meiotic prophase in the C. elegans germ line. Dev. Biol. 308, 206–221. Abstract Article
Jaramillo-Lambert, A., Harigaya, Y., Vitt, J., Villeneuve, A., and Engebrecht, J. (2010). Meiotic errors activate checkpoints that improve gamete quality without triggering apoptosis in male germ cells. Curr. Biol. 20, 2078–2089. Abstract Article
Kaitna, S., Pasierbek, P., Jantsch, M., Loidl, J., and Glotzer, M. (2002). The aurora B kinase AIR-2 regulates kinetochores during mitosis and is required for separation of homologous chromosomes during meiosis. Curr. Biol. 12, 798–812. Abstract Article
Kauppi, L., Barchi, M., Lange, J., Baudat, F., Jasin, M., and Keeney, S. (2013). Numerical constraints and feedback control of double-strand breaks in mouse meiosis. Genes Dev. 27, 873–886. Abstract Article
Kaur, T., and Rockman, M. V (2014). Crossover heterogeneity in the absence of hotspots in Caenorhabditis elegans. Genetics 196, 137–148. Abstract Article
Kelly, K.O., Dernburg, A.F., Stanfield, G.M., and Villeneuve, A.M. (2000). Caenorhabditis elegans msh-5 is required for both normal and radiation-induced meiotic crossing over but not for completion of meiosis. Genetics 156, 617–630. Abstract
Kelly, W.G., Schaner, C.E., Dernburg, A.F., Lee, M.-H., Kim, S.K., Villeneuve, A.M., and Reinke, V. (2002). X-chromosome silencing in the germline of C. elegans. Development 129, 479–492. Abstract
Kim, S., Spike, C., and Greenstein, D. (2013). Control of oocyte growth and meiotic maturation in Caenorhabditis elegans. Adv. Exp. Med. Biol. 757, 277–320. Abstract Article
Kim, Y., Rosenberg, S.C., Kugel, C.L., Kostow, N., Rog, O., Davydov, V., Su, T.Y., Dernburg, A.F., and Corbett, K.D. (2014). The chromosome axis controls meiotic events through a hierarchical assembly of HORMA domain proteins. Dev. Cell 31, 487–502. Abstract Article
Kimble, J. and Crittenden, S.L. Germline proliferation and its control (August 15, 2005), WormBook, ed. The C. elegans Research Community, WormBook, doi/10.1895/wormbook.1.13.1,http://www.wormbook.org. Abstract
Kohl, K.P., and Sekelsky, J. (2013). Meiotic and mitotic recombination in meiosis. Genetics 194, 327–334. Abstract Article
Labella, S., Woglar, A., Jantsch, V., and Zetka, M. (2011). Polo kinases establish links between meiotic chromosomes and cytoskeletal forces essential for homolog pairing. Dev. Cell 21, 948–958. Abstract Article
Labrador, L., Barroso, C., Lightfoot, J., Müller-Reichert, T., Flibotte, S., Taylor, J., Moerman, D.G., Villeneuve, A.M., and Martinez-Perez, E. (2013). Chromosome movements promoted by the mitochondrial protein SPD-3 are required for homology search during Caenorhabditis elegans meiosis. PLoS Genet. 9, e1003497. Abstract Article
Lackner, M.R., and Kim, S.K. (1998). Genetic analysis of the Caenorhabditis elegans MAP kinase gene mpk-1. Genetics 150, 103–117. Abstract
Lamelza, P., and Bhalla, N. (2012). Histone methyltransferases MES-4 and MET-1 promote meiotic checkpoint activation in Caenorhabditis elegans. PLoS Genet. 8, e1003089. Abstract Article
Lange, J., Pan, J., Cole, F., Thelen, M.P., Jasin, M., and Keeney, S. (2011). ATM controls meiotic double-strand-break formation. Nature 479, 237–240. Abstract Article
Lee, M.-H., Ohmachi, M., Arur, S., Nayak, S., Francis, R., Church, D., Lambie, E., and Schedl, T. (2007). Multiple functions and dynamic activation of MPK-1 extracellular signal-regulated kinase signaling in Caenorhabditis elegans germline development. Genetics 177, 2039–2062. Abstract Article
Lemmens, B.B., Johnson, N.M., and Tijsterman, M. (2013). COM-1 promotes homologous recombination during Caenorhabditis elegans meiosis by antagonizing Ku-mediated non-homologous end joining. PLoS Genet. 9, e1003276. Abstract Article
Libuda, D.E., Uzawa, S., Meyer, B.J., and Villeneuve, A.M. (2013). Meiotic chromosome structures constrain and respond to designation of crossover sites. Nature 502, 703–706. Abstract Article
Lightfoot, J., Testori, S., Barroso, C., and Martinez-Perez, E. (2011). Loading of meiotic cohesin by SCC-2 is required for early processing of DSBs and for the DNA damage checkpoint. Curr. Biol. 21, 1421–1430. Abstract Article
Lim, J.G.Y., Stine, R.R.W., and Yanowitz, J.L. (2008). Domain-specific regulation of recombination in Caenorhabditis elegans in response to temperature, age and sex. Genetics 180, 715–726. Abstract Article
MacQueen, A.J., Colaiácovo, M.P., McDonald, K., and Villeneuve, A.M. (2002). Synapsis-dependent and -independent mechanisms stabilize homolog pairing during meiotic prophase in C. elegans. Genes Dev. 16, 2428–2442. Abstract Article
MacQueen, A.J., Phillips, C.M., Bhalla, N., Weiser, P., Villeneuve, A.M., and Dernburg, A.F. (2005). Chromosome sites play dual roles to establish homologous synapsis during meiosis in C. elegans. Cell 123, 1037–1050. Abstract Article
MacQueen, A.J., and Villeneuve, A.M. (2001). Nuclear reorganization and homologous chromosome pairing during meiotic prophase require C. elegans chk-2. Genes Dev. 15, 1674–1687. Abstract Article
Mains, P.E., Kemphues, K.J., Sprunger, S.A., Sulston, I.A., and Wood, W.B. (1990). Mutations affecting the meiotic and mitotic divisions of the early Caenorhabditis elegans embryo. Genetics 126, 593–605. Abstract
Malone, C.J., Misner, L., Le Bot, N., Tsai, M.-C., Campbell, J.M., Ahringer, J., and White, J.G. (2003). The C. elegans hook protein, ZYG-12, mediates the essential attachment between the centrosome and nucleus. Cell 115, 825–836. Abstract Article
Martin, J.S., Winkelmann, N., Petalcorin, M.I.R., McIlwraith, M.J., and Boulton, S.J. (2005). RAD-51-dependent and -independent roles of a Caenorhabditis elegans BRCA2-related protein during DNA double-strand break repair. Mol. Cell. Biol. 25, 3127–3139. Abstract Article
Martinez-Perez, E., Schvarzstein, M., Barroso, C., Lightfoot, J., Dernburg, A.F., and Villeneuve, A.M. (2008). Crossovers trigger a remodeling of meiotic chromosome axis composition that is linked to two-step loss of sister chromatid cohesion. Genes Dev. 22, 2886–2901. Abstract
Martinez-Perez, E., and Villeneuve, A.M. (2005). HTP-1-dependent constraints coordinate homolog pairing and synapsis and promote chiasma formation during C. elegans meiosis. Genes Dev. 19, 2727–2743. Abstract Article
McKim, K.S., Howell, A.M., and Rose, A.M. (1988). The effects of translocations on recombination frequency in Caenorhabditis elegans. Genetics 120, 987–1001. Abstract
McNicoll, F., Stevense, M., and Jessberger, R. (2013). Cohesin in gametogenesis. Curr. Top. Dev. Biol. 102, 1–34. Abstract Article
Meneely, P.M., Farago, A.F., and Kauffman, T.M. (2002). Crossover distribution and high interference for both the X chromosome and an autosome during oogenesis and spermatogenesis in Caenorhabditis elegans. Genetics 162, 1169–1177. Abstract
Meneely, P.M., McGovern, O.L., Heinis, F.I., and Yanowitz, J.L. (2012). Crossover distribution and frequency are regulated by him-5 in Caenorhabditis elegans. Genetics 190, 1251–1266. Abstract Article
Mets, D.G., and Meyer, B.J. (2009). Condensins regulate meiotic DNA break distribution, thus crossover frequency, by controlling chromosome structure. Cell 139, 73–86. Abstract Article
Meyer, B. J. X-Chromosome dosage compensation (June 25, 2005), WormBook, ed. The C. elegans Research Community, WormBook, doi/10.1895/wormbook.1.8.1, http://www.wormbook.org.
Meyer, B.J. (2010). Targeting X chromosomes for repression. Curr. Opin. Genet. Dev. 20, 179–189. Abstract Article
Minn, I.L., Rolls, M.M., Hanna-Rose, W., and Malone, C.J. (2009). SUN-1 and ZYG-12, mediators of centrosome-nucleus attachment, are a functional SUN/KASH pair in Caenorhabditis elegans. Mol. Biol. Cell 20, 4586–4595. Abstract Article
Mlynarczyk-Evans, S., Roelens, B., and Villeneuve, A.M. (2013). Evidence that masking of synapsis imperfections counterbalances quality control to promote efficient meiosis. PLoS Genet. 9, e1003963. Abstract Article
Monen, J., Maddox, P.S., Hyndman, F., Oegema, K., and Desai, A. (2005). Differential role of CENP-A in the segregation of holocentric C. elegans chromosomes during meiosis and mitosis. Nat. Cell Biol. 7, 1248–1255. Abstract Article
Muller, H.J. (1916). The mechanism of crossing-over. Am. Nat. 50, 193–221, 284–305, 350–366, 421–434. Article
Muscat, C.C., Torre-Santiago, K.M., Tran, M.V., Powers, J.A., and Wignall, S.M. (2015). Kinetochore-independent chromosome segregation driven by lateral microtubule bundles. eLife 4, e06462. Abstract Article
Nabeshima, K., Mlynarczyk-Evans, S., and Villeneuve, A.M. (2011). Chromosome painting reveals asynaptic full alignment of homologs and HIM-8-dependent remodeling of X chromosome territories during Caenorhabditis elegans meiosis. PLoS Genet. 7, e1002231. Abstract Article
Nabeshima, K., Villeneuve, A.M., and Colaiácovo, M.P. (2005). Crossing over is coupled to late meiotic prophase bivalent differentiation through asymmetric disassembly of the SC. J. Cell Biol. 168, 683–689. Abstract
Nabeshima, K., Villeneuve, A.M., and Hillers, K.J. (2004). Chromosome-wide regulation of meiotic crossover formation in Caenorhabditis elegans requires properly assembled chromosome axes. Genetics 168, 1275–1292. Abstract
Nicolas, A., Treco, D., Schultes, N.P., and Szostak, J.W. (1989). An initiation site for meiotic gene conversion in the yeast Saccharomyces cerevisiae. Nature 338, 35–39. Abstract
Nottke, A.C., Beese-Sims, S.E., Pantalena, L.F., Reinke, V., Shi, Y., and Colaiácovo, M.P. (2011). SPR-5 is a histone H3K4 demethylase with a role in meiotic double-strand break repair. Proc. Natl. Acad. Sci. U. S. A. 108, 12805–12810. Abstract
O'Neil, N.J., Martin, J.S., Youds, J.L., Ward, J.D., Petalcorin, M.I.R., Rose, A.M., and Boulton, S.J. (2013). Joint molecule resolution requires the redundant activities of MUS-81 and XPF-1 during Caenorhabditis elegans meiosis. PLoS Genet. 9, e1003582. Abstract
Pâques, F., and Haber, J.E. (1999). Multiple pathways of recombination induced by double-strand breaks in Saccharomyces cerevisiae. Microbiol. Mol. Biol. Rev. 63, 349–404. Abstract
Pasierbek, P., Födermayr, M., Jantsch, V., Jantsch, M., Schweizer, D., and Loidl, J. (2003). The Caenorhabditis elegans SCC-3 homologue is required for meiotic synapsis and for proper chromosome disjunction in mitosis and meiosis. Exp. Cell Res. 289, 245–255. Abstract Article
Pasierbek, P., Jantsch, M., Melcher, M., Schleiffer, A., Schweizer, D., and Loidl, J. (2001). A Caenorhabditis elegans cohesion protein with functions in meiotic chromosome pairing and disjunction. Genes Dev. 15, 1349–1360. Abstract Article
Penkner, A., Portik-Dobos, Z., Tang, L., Schnabel, R., Novatchkova, M., Jantsch, V., and Loidl, J. (2007a). A conserved function for a Caenorhabditis elegans Com1/Sae2/CtIP protein homolog in meiotic recombination. EMBO J. 26, 5071–5082. Abstract Article
Penkner, A., Tang, L., Novatchkova, M., Ladurner, M., Fridkin, A., Gruenbaum, Y., Schweizer, D., Loidl, J., and Jantsch, V. (2007b). The nuclear envelope protein Matefin/SUN-1 is required for homologous pairing in C. elegans meiosis. Dev. Cell 12, 873–885. Abstract Article
Penkner, A.M., Fridkin, A., Gloggnitzer, J., Baudrimont, A., Machacek, T., Woglar, A., Csaszar, E., Pasierbek, P., Ammerer, G., Gruenbaum, Y., and Jantsch, V.l. (2009). Meiotic chromosome homology search involves modifications of the nuclear envelope protein Matefin/SUN-1. Cell 139, 920–933. Abstract
Petalcorin, M.I.R., Galkin, V.E., Yu, X., Egelman, E.H., and Boulton, S.J. (2007). Stabilization of RAD-51-DNA filaments via an interaction domain in Caenorhabditis elegans BRCA2. Proc. Natl. Acad. Sci. U. S. A. 104, 8299–8304. Abstract Article
Petalcorin, M.I.R., Sandall, J., Wigley, D.B., and Boulton, S.J. (2006). CeBRC-2 stimulates D-loop formation by RAD-51 and promotes DNA single-strand annealing. J. Mol. Biol. 361, 231–242. Abstract
Phillips, C.M., and Dernburg, A.F. (2006). A family of zinc-finger proteins is required for chromosome-specific pairing and synapsis during meiosis in C. elegans. Dev. Cell 11, 817–829. Abstract
Phillips, C.M., Meng, X., Zhang, L., Chretien, J.H., Urnov, F.D., and Dernburg, A.F. (2009). Identification of chromosome sequence motifs that mediate meiotic pairing and synapsis in C. elegans. Nat. Cell Biol. 11, 934–942. Abstract
Phillips, C.M., Wong, C., Bhalla, N., Carlton, P.M., Weiser, P., Meneely, P.M., and Dernberg, A.F. (2005). HIM-8 binds to the X chromosome pairing center and mediates chromosome-specific meiotic synapsis. Cell 123, 1051–1063. Abstract
Qiao, H., Prasada Rao, H.B.D., Yang, Y., Fong, J.H., Cloutier, J.M., Deacon, D.C., et al. (2014). Antagonistic roles of ubiquitin ligase HEI10 and SUMO ligase RNF212 regulate meiotic recombination. Nat. Genet. 46, 194–199. Article
Reddy, K.C., and Villeneuve, A.M. (2004). C. elegans HIM-17 links chromatin modification and competence for initiation of meiotic recombination. Cell 118, 439–452. Abstract Article
Rinaldo, C., Bazzicalupo, P., Ederle, S., Hilliard, M., and La Volpe, A. (2002). Roles for Caenorhabditis elegans rad-51 in meiosis and in resistance to ionizing radiation during development. Genetics 160, 471–479. Abstract
Robert, V.J., Davis, M.W., Jorgensen, E.M., and Bessereau, J.-L. (2008). Gene conversion and end-joining-repair double-strand breaks in the Caenorhabditis elegans germline. Genetics 180, 673–679. Abstract Article
Rockman, M.V., and Kruglyak, L. (2009). Recombinational landscape and population genomics of Caenorhabditis elegans. PLoS Genet. 5, e1000419. Abstract Article
Rog, O., and Dernburg, A.F. (2013). Chromosome pairing and synapsis during Caenorhabditis elegans meiosis. Curr. Opin. Cell Biol. 25, 349–56. Abstract
Rog, O., and Dernburg, A.F. (2015). Direct visualization reveals kinetics of meiotic chromosome synapsis. Cell Rep. 10, 1639-1645. Abstract Article
Rogers, E., Bishop, J.D., Waddle, J.A., Schumacher, J.M., and Lin, R. (2002). The aurora kinase AIR-2 functions in the release of chromosome cohesion in Caenorhabditis elegans meiosis. J. Cell Biol. 157, 219–229. Abstract Article
Rose, A.M., and Baillie, D.L. (1979a). A mutation in Caenorhabditis elegans that increases recombination frequency more than threefold. Nature 281, 599–600. Abstract Article
Rose, A.M., and Baillie, D.L. (1979b). The effect of temperature and parental age on recombination and nondisjunction in Caenorhabditis elegans. Genetics 92, 409–418. Abstract
Rose, A.M., Baillie, D.L., and Curran, J. (1984). Meiotic pairing behavior of two free duplications of linkage group I in Caenorhabditis elegans. Mol. Gen. Genet. 195, 52–56. Abstract Article
Rosenbluth, R.E., and Baillie, D.L. (1981). The genetic analysis of a reciprocal translocation, eT1(III; V), in Caenorhabditis elegans. Genetics 99, 415–428. Abstract
Rosu, S., Libuda, D.E., and Villeneuve, A.M. (2011). Robust crossover assurance and regulated interhomolog access maintain meiotic crossover number. Science 334, 1286–1289. Abstract Article
Rosu, S., Zawadzki, K.A., Stamper, E.L., Libuda, D.E., Reese, A.L., Dernburg, A.F., and Villeneuve, A.M. (2013). The C. elegans DSB-2 protein reveals a regulatory network that controls competence for meiotic DSB formation and promotes crossover assurance. PLoS Genet. 9, e1003674. Abstract Article
Saito, T.T., Lui, D.Y., Kim, H.-M., Meyer, K., and Colaiácovo, M.P. (2013). Interplay between structure-specific endonucleases for crossover control during Caenorhabditis elegans meiosis. PLoS Genet. 9, e1003586. Abstract Article
Saito, T.T., Mohideen, F., Meyer, K., Harper, J.W., and Colaiácovo, M.P. (2012). SLX-1 is required for maintaining genomic integrity and promoting meiotic noncrossovers in the Caenorhabditis elegans germline. PLoS Genet. 8, e1002888. Abstract Article
Saito, T.T., Youds, J.L., Boulton, S.J., and Colaiácovo, M.P. (2009). Caenorhabditis elegans HIM-18/SLX-4 interacts with SLX-1 and XPF-1 and maintains genomic integrity in the germline by processing recombination intermediates. PLoS Genet. 5, e1000735. Abstract Article
Sanford, C., and Perry, M.D. (2001). Asymmetrically distributed oligonucleotide repeats in the Caenorhabditis elegans genome sequence that map to regions important for meiotic chromosome segregation. Nucleic Acids Res. 29, 2920–2926. Abstract Article
Sato, A., Isaac, B., Phillips, C.M., Rillo, R., Carlton, P.M., Wynne, D.J., Kasad, R.A., and Dernburg. A.F. (2009). Cytoskeletal forces span the nuclear envelope to coordinate meiotic chromosome pairing and synapsis. Cell 139, 907–919. Abstract Article
Sato-Carlton, A., Li, X., Crawley, O., Testori, S., Martinez-Perez, E., Sugimoto, A., and Carlton, P.M. (2014). Protein phosphatase 4 promotes chromosome pairing and Ssynapsis, and contributes to maintaining crossover competence with increasing age. PLoS Genet. 10, e1004638. Abstract Article
Schild-Prüfert, K., Saito, T.T., Smolikov, S., Gu, Y., Hincapie, M., Hill, D.E., Vidal, M., McDonald, K., and Colaiácovo, M.P. (2011). Organization of the synaptonemal complex during meiosis in Caenorhabditis elegans. Genetics 189, 411–421. Abstract Article
Schumacher, B., Hofmann, K., Boulton, S., and Gartner, A. (2001). The C. elegans homolog of the p53 tumor suppressor is required for DNA damage-induced apoptosis. Curr. Biol. 11, 1722–1727. Abstract Article
Schvarzstein, M., Pattabiraman, D., Bembenek, J.N., and Villeneuve, A.M. (2013). Meiotic HORMA domain proteins prevent untimely centriole disengagement during Caenorhabditis elegans spermatocyte meiosis. Proc. Natl. Acad. Sci. U. S. A. 110, E898–E907. Abstract Article
Schvarzstein, M., Pattabiraman, D., Libuda, D.E., Ramadugu, A., Tam, A., Martinez-Perez, E., Roelens, B., Zawadzki, K.A., Yokoo, R., Rosu, S., et al. (2014). DNA helicase HIM-6/BLM both promotes MutSγ-dependent crossovers and antagonizes MutSγ-independent interhomolog associations during Caenorhabditis elegans meiosis. Genetics 198, 193–207. Abstract Article
Schvarzstein, M., Wignall, S.M., and Villeneuve, A.M. (2010). Coordinating cohesion, co-orientation, and congression during meiosis: lessons from holocentric chromosomes. Genes Dev. 24, 219–228. Abstract Article
Segbert, C., Barkus, R., Powers, J., Strome, S., Saxton, W.M., and Bossinger, O. (2003). KLP-18, a Klp2 kinesin, is required for assembly of acentrosomal meiotic spindles in Caenorhabditis elegans. Mol. Biol. Cell 14, 4458–4469. Abstract Article
Severson, A.F., Ling, L., Zuylen, V. van, and Meyer, B.J. (2009). The axial element protein HTP-3 promotes cohesin loading and meiotic axis assembly in C. elegans to implement the meiotic program of chromosome segregation. Genes Dev. 23, 1763–1778. Abstract Article
Severson, A.F., and Meyer, B.J. (2014). Divergent kleisin subunits of cohesin specify mechanisms to tether and release meiotic chromosomes. eLife 3, e03467. Abstract Article
Shakes, D.C., Wu, J.-C., Sadler, P.L., Laprade, K., Moore, L.L., Noritake, A., and Chu, D.S. (2009). Spermatogenesis-specific features of the meiotic program in Caenorhabditis elegans. PLoS Genet. 5, e1000611. Abstract Article
Silva, N., Adamo, A., Santonicola, P., Martinez-Perez, E., and La Volpe, A. (2013). Pro-crossover factors regulate damage-dependent apoptosis in the Caenorhabditis elegans germ line. Cell Death Differ. 20, 1209–1218. Abstract Article
Silva, N., Ferrandiz, N., Barroso, C., Tognetti, S., Lightfoot, J., Telecan, O., Encheva, V., Faull, P., Hanni, S., Furger, A., et al. (2014). The fidelity of synaptonemal complex assembly is regulated by a signaling mechanism that controls early meiotic progression. Dev. Cell 31, 503–511. Abstract
Siomos, M.F., Badrinath, A., Pasierbek, P., Livingstone, D., White, J., Glotzer, M., and Nasmyth, K. (2001). Separase is required for chromosome segregation during meiosis I in Caenorhabditis elegans. Curr. Biol. 11, 1825–1835. Abstract Article
Smolikov, S., Eizinger, A., Hurlburt, A., Rogers, E., Villeneuve, A.M., and Colaiácovo, M.P. (2007a). Synapsis-defective mutants reveal a correlation between chromosome conformation and the mode of double-strand break repair during Caenorhabditis elegans meiosis. Genetics 176, 2027–2033. Abstract Article
Smolikov, S., Eizinger, A., Schild-Prufert, K., Hurlburt, A., McDonald, K., Engebrecht, J., et al. (2007b). SYP-3 restricts synaptonemal complex assembly to bridge paired chromosome axes during meiosis in Caenorhabditis elegans. Genetics 176, 2015–2025. Abstract Article
Smolikov, S., Schild-Prüfert, K., and Colaiácovo, M.P. (2008). CRA-1 uncovers a double-strand break-dependent pathway promoting the assembly of central region proteins on chromosome axes during C. elegans meiosis. PLoS Genet. 4, e1000088. Abstract Article
Smolikov, S., Schild-Prüfert, K., and Colaiácovo, M.P. (2009). A yeast two-hybrid screen for SYP-3 interactors identifies SYP-4, a component required for synaptonemal complex assembly and chiasma formation in Caenorhabditis elegans meiosis. PLoS Genet. 5, e1000669. Abstract Article
Song, A., Labella, S., Korneeva, N.L., Keiper, B.D., Aamodt, E.J., Zetka, M., and Rhoads, R.E. (2010). A C. elegans eIF4E-family member upregulates translation at elevated temperatures of mRNAs encoding MSH-5 and other meiotic crossover proteins. J. Cell Sci. 123, 2228–2237. Abstract Article
Spike, C.A., Coetzee, D., Eichten, C., Wang, X., Hansen, D., and Greenstein, D. (2014). The TRIM-NHL protein LIN-41 and the OMA RNA-binding proteins antagonistically control the prophase-to-metaphase transition and growth of Caenorhabditis elegans oocytes. Genetics 198, 1535–1558. Abstract Article
Srayko, M., Buster, D.W., Bazirgan, O.A., McNally, F.J., and Mains, P.E. (2000). MEI-1/MEI-2 katanin-like microtubule severing activity is required for Caenorhabditis elegans meiosis. Genes Dev. 14, 1072–1084. Abstract
Stamper, E.L., Rodenbusch, S.E., Rosu, S., Ahringer, J., Villeneuve, A.M., and Dernburg, A.F. (2013). Identification of DSB-1, a protein required for initiation of meiotic recombination in Caenorhabditis elegans, illuminates a crossover assurance checkpoint. PLoS Genet. 9, e1003679. Abstract Article
Stevens, D., Oegema, K., and Desai, A. (2013). Meiotic double-strand breaks uncover and protect against mitotic errors in the C. elegans germline. Curr. Biol. 23, 2400–2406. Abstract Article
Takanami, T., Mori, A., Takahashi, H., Horiuchi, S., and Higashitani, A. (2003). Caenorhabditis elegans Ce-rdh-1/rad-51 functions after double-strand break formation of meiotic recombination. Chromosome Res. 11, 125–135. Abstract Article
Tang, L., Machacek, T., Mamnun, Y.M., Penkner, A., Gloggnitzer, J., Wegrostek, C., Konrat, R., Jantsch, M.F., Loidl, J., and Jantsch, V. (2010). Mutations in Caenorhabditis elegans him-19 show meiotic defects that worsen with age. Mol. Biol. Cell 21, 885–896. Abstract
Thacker, D., Mohibullah, N., Zhu, X., and Keeney, S. (2014). Homologue engagement controls meiotic DNA break number and distribution. Nature 510, 241–246. Abstract Article
Tsai, C.J., Mets, D.G., Albrecht, M.R., Nix, P., Chan, A., and Meyer, B.J. (2008). Meiotic crossover number and distribution are regulated by a dosage compensation protein that resembles a condensin subunit. Genes Dev. 22, 194–211. Abstract Article
Tzur, Y.B., de Carvalho, C.E., Nadarajan, S., Bostelen, I. Van, Gu, Y., Chu, D.S., Cheeseman, I.M., Colaiácovo, M.P. (2012). LAB-1 targets PP1 and restricts Aurora B kinase upon entrance into meiosis to promote sister chromatid cohesion. PLoS Biol. 10, e1001378. Abstract Article
Villeneuve, A.M. (1994). A cis-acting locus that promotes crossing over between X chromosomes in Caenorhabditis elegans. Genetics 136, 887–902. Abstract
van der Voet, M., Berends, C.W.H., Perreault, A., Nguyen-Ngoc, T., Gönczy, P., Vidal, M., Boxem, M., and van den Heuvel, S. (2009). NuMA-related LIN-5, ASPM-1, calmodulin and dynein promote meiotic spindle rotation independently of cortical LIN-5/GPR/Gα. Nat. Cell Biol. 11, 269–277. Abstract Article
Wagner, C.R., Kuervers, L., Baillie, D.L., and Yanowitz, J.L. (2010). xnd-1 regulates the global recombination landscape in Caenorhabditis elegans. Nature 467, 839–843. Abstract Article
Wang, F., Yoder, J., Antoshechkin, I., and Han, M. (2003). Caenorhabditis elegans EVL-14/PDS-5 and SCC-3 are essential for sister chromatid cohesion in meiosis and mitosis. Mol. Cell. Biol. 23, 7698–7707. Abstract Article
Ward, J.D., Muzzini, D.M., Petalcorin, M.I.R., Martinez-Perez, E., Martin, J.S., Plevani, P., Cassata, G., Marini, F., and Boulton, S.J. (2010). Overlapping mechanisms promote postsynaptic RAD-51 filament disassembly during meiotic double-strand break repair. Mol. Cell 37, 259–272. Abstract Article
Wignall, S.M., and Villeneuve, A.M. (2009). Lateral microtubule bundles promote chromosome alignment during acentrosomal oocyte meiosis. Nat. Cell Biol. 11, 839–844. Abstract Article
Winand, N.J., Panzer, J.A., and Kolodner, R.D. (1998). Cloning and characterization of the human and Caenorhabditis elegans homologs of the Saccharomyces cerevisiae MSH5 gene. Genomics 53, 69–80. Abstract Article
Woglar, A., Daryabeigi, A., Adamo, A., Habacher, C., Machacek, T., La Volpe, A., and Jantsch, V. (2013). Matefin/SUN-1 phosphorylation is part of a surveillance mechanism to coordinate chromosome synapsis and recombination with meiotic progression and chromosome movement. PLoS Genet. 9, e1003335. Abstract Article
Wood, A.J., Severson, A.F., and Meyer, B.J. (2010). Condensin and cohesin complexity: the expanding repertoire of functions. Nat. Rev. Genet. 11, 391–404. Abstract Article
Wynne, D.J., Rog, O., Carlton, P.M., and Dernburg, A.F. (2012). Dynein-dependent processive chromosome motions promote homologous pairing in C. elegans meiosis. J. Cell Biol. 196, 47–64. Abstract Article
Yang, H., Mains, P.E., and McNally, F.J. (2005). Kinesin-1 mediates translocation of the meiotic spindle to the oocyte cortex through KCA-1, a novel cargo adapter. J. Cell Biol. 169, 447–457. Abstract Article
Yang, H., McNally, K., and McNally, F.J. (2003). MEI-1/katanin is required for translocation of the meiosis I spindle to the oocyte cortex in C elegans. Dev. Biol. 260, 245–59. Abstract Article
Yin, Y., and Smolikove, S. (2013). Impaired resection of meiotic double-strand breaks channels repair to nonhomologous end joining in Caenorhabditis elegans. Mol. Cell. Biol. 33, 2732–2747. Abstract Article
Yokoo, R., Zawadzki, K.A., Nabeshima, K., Drake, M., Arur, S., and Villeneuve, A.M. (2012). COSA-1 reveals robust homeostasis and separable licensing and reinforcement steps governing meiotic crossovers. Cell 149, 75–87. Abstract Article
Youds, J.L., Mets, D.G., McIlwraith, M.J., Martin, J.S., Ward, J.D., ONeil, N.J., Rose, A.M., West, S.C., Meyer, B.J., and Boulton, S.J. (2010). RTEL-1 enforces meiotic crossover interference and homeostasis. Science 327, 1254–1258. Abstract Article
Zalevsky, J., MacQueen, A.J., Duffy, J.B., Kemphues, K.J., and Villeneuve, A.M. (1999). Crossing over during Caenorhabditis elegans meiosis requires a conserved MutS-based pathway that is partially dispensable in budding yeast. Genetics 153, 1271–1283. Abstract
Zetka, M.C., Kawasaki, I., Strome, S., and Muller, F. (1999). Synapsis and chiasma formation in Caenorhabditis elegans require HIM-3, a meiotic chromosome core component that functions in chromosome segregation. Genes Dev. 13, 2258–2270. Abstract Article
Zetka, M.C., and Rose, A.M. (1990). Sex-related differences in crossing over in Caenorhabditis elegans. Genetics 126, 355–363. Abstract
Zetka, M.C., and Rose, A.M. (1995). Mutant rec-1 eliminates the meiotic pattern of crossing over in Caenorhabditis elegans. Genetics 141, 1339–1349. Abstract
Zhang, W., Miley, N., Zastrow, M.S., MacQueen, A.J., Sato, A., Nabeshima, K., Martinez-Perez, E., Mlynarczyk-Evans, S., Carlton, P.M., and Villeneuve, A.M. (2012). HAL-2 promotes homologous pairing during Caenorhabditis elegans meiosis by antagonizing inhibitory effects of synaptonemal complex precursors. PLoS Genet. 8, e1002880. Abstract Article
*Edited by Anne Villeneuve and David Greenstein Last revised May 26, 2015. Published in its final form May 4, 2017. This chapter should be cited as: Hillers K.J., Jantsch V., Martinez-Perez E., Yanowitz J.L. Meiosis. (May 4, 2017), WormBook, ed. The C. elegans Research Community, WormBook, doi/10.1895/wormbook.1.178.1, http://www.wormbook.org.
Copyright: © 2016 Kenneth J Hillers, Verena Jantsch, Enrique Martinez-Perez, Judith L Yanowitz. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
§To whom correspondence should be addressed. Email: [email protected]; [email protected]; [email protected]; [email protected]
 All WormBook content, except where otherwise noted, is licensed under a Creative Commons Attribution License.
All WormBook content, except where otherwise noted, is licensed under a Creative Commons Attribution License.