Embryo series courtesy of Einhard Schierenberg
Embryo series courtesy of Einhard SchierenbergTable of Contents
Abstract
C. elegans represents a favorable system to study the extraordinarily complicated process of eukaryotic protein synthesis, which involves over 100 RNAs and over 200 polypeptides just for the core machinery. Initial research in protein synthesis relied on fractionated mammalian and plant systems, but in the mid-1970s, the powerful genetics of Saccharomyces cerevisiae began to yield new insights for translation in all eukaryotes. C. elegans has many features of higher eukaryotes that are not shared by yeast. This allows protein synthesis researchers to combine biochemistry, cell biology, developmental biology, genetics, and genomics to study regulation of gene expression at the translational level. Most components of the core translational machinery have been identified in C. elegans, including rRNAs, 5S RNA, tRNAs, ribosomal proteins, and aminoacyl tRNA synthetases. C. elegans has amino acid sequence homologs for 56 of the known initiation, elongation, and release factor polypeptides, but few of these have been isolated, functionally identified, or studied at the biochemical level. Similarly, C. elegans has homologs for 22 components of the major signal transduction pathways implicated in control of protein synthesis. The translational efficiency of individual mRNAs relies on cis-regulatory elements that include either a 7-methylguanosine- or 2,2,7-trimethylguanosine-containing cap, the 5'-terminal spliced leader, sequence elements in the 3'-untranslated regions, and the 3'-terminal poly(A) tract. Several key developmental pathways in C. elegans are predominantly governed by translational mechanisms. Some evidence has been presented that well described regulatory mechanisms in other organisms, including covalent modification of translation factors, sequestration of translation factors, and mRNA-specific changes in poly(A) length, also occur in C. elegans. The most interesting unexplored questions may involve changes in the translation of individual mRNAs during development, in response to physiological changes, or after genetic manipulations. Given the highly developed state of C. elegans genomics, it can be expected that future application of computational tools, including data visualization, will help detect new instances of translational control.
Although "gene expression" is often used synonymously with "transcription", the steady-state levels of proteins in eukaryotic cells are also strongly dependent on translational regulatory mechanisms. Two very different types of translational control occur: global and mRNA-specific. Global control generally involves alterations in the levels, intrinsic activities, or availability of translation factors, whereas mRNA-specific control involves proteins or RNAs that interact with a subset of mRNAs. The overall rate of protein synthesis as well as the translational efficiencies of individual mRNAs are regulated in response to nutritional, hormonal, cellular stress, and developmental signals (Sonenberg et al., 2000).
C. elegans represents an especially favorable system to study the extraordinarily complicated process of eukaryotic protein synthesis, which involves over 100 RNAs and over 200 polypeptides just for the core machinery. The first decades of research in eukaryotic protein synthesis relied on fractionated mammalian and plant systems, with little or no input of genetics. This began to change in the 1970's when the powerful genetics of Saccharomyces cerevisiae was brought to bear on central questions in protein synthesis. From this research came important new insights for translation in all eukaryotes, including discovery of the initiation codon scanning mechanism (Sherman and Stewart, 1975), new protein synthesis factors and regulatory kinases (Hinnebusch, 1997), previously unknown interactions among initiation factors (Asano et al., 2000), the core structure of eIF3, the most complex of the initiation factors (Phan et al., 1998), and new regulatory pathways for the control of protein synthesis (Hinnebusch and Fink, 1983). C. elegans, however, has many features of higher eukaryotes that are not shared by yeast, e.g., tissues, organs, muscles, a nervous system, developmental stages, cell lineages, etc., which involve processes regulated at the translational level. Furthermore, signaling pathways leading to protein synthesis are considerably more similar between C. elegans and humans than between yeast and humans. Thus, C. elegans allows protein synthesis researchers to combine biochemistry, cell biology, genetics, and genomics to understand fundamental questions about the regulation of gene expression at the translational level.
From studies in mammals, yeasts, and plants, it is known that the three steps of protein synthesis are catalyzed by three groups of proteins: initiation, elongation, and release factors (Hershey and Merrick, 2000). A different class of initiation factors (eIF1, eIF2, etc.) catalyzes each step of initiation (Figure 1). [A uniform nomenclature system for translation factors is used here (Clark et al., 1996)]. A ternary complex of eIF2•GTP•Met-tRNAi binds to the 40S ribosomal subunit to form the 43S initiation complex. Recruitment of mRNA to the 43S initiation complex to form the 48S initiation complex requires eIF3, the poly(A)-binding protein (PABP), and the eIF4 proteins. eIF3 is a ~800-kDa multimer that is also required for Met-tRNAi binding to the 40S subunit (molecular masses refer to the mammalian factors). PABP is a 70-kDa protein that specifically binds poly(A) and homo-oligomerizes. The eIF4 factors consist of: eIF4A, a 46-kDa RNA helicase; eIF4B, a 70-kDa RNA-binding and RNA-annealing protein; eIF4H, a 25-kDa protein that acts with eIF4B to stimulate eIF4A helicase activity; eIF4E, a 25-kDa cap-binding protein; and eIF4G, a 185-kDa protein that specifically binds to and co-localizes all of the other proteins involved in mRNA recruitment on the 40S subunit.
The 48S complex consists of the eIF4 factors plus PABP, eIF1, eIF1A, eIF3, eIF5, and the eIF2•GTP•Met-tRNAi ternary complex bound to the 40S subunit (Figure 2). It scans until the first AUG in good sequence context is encountered. Scanning requires ATP hydrolysis by eIF4A and the presence of eIF1 and eIF1A. Then eIF5 and eIF5B stimulate GTP hydrolysis by eIF2, followed by 60S joining to form the 80S complex. The released eIF2•GDP is recycled to eIF2•GTP by the guanine nucleotide exchange factor eIF2B. The first elongator aminoacyl-tRNA is brought to the ribosomal A-site by eEF1, after which the first peptide bond is formed. This is followed by a cycle of GTP hydrolysis and exchange. Translocation is catalyzed by eEF2 with another cycle of GTP hydrolysis and exchange. When the ribosome reaches a termination codon, the release factor eRF1 catalyzes termination, and then the GTPase eRF3 ejects eRF1 from the ribosome.
|
Figure 2. Model for the 48S initiation complex. The interactions among eIF1, eIF2, eIF3, eIF4A, eIF4E, eIF4G, eIF5, Mnk, PABP, mRNA, and the 40S ribosomal subunit are shown. The thin line represents mRNA, with the wavy line indicating mRNA secondary structure. Meti is the initiator tRNA. The sizes of protein depictions are roughly proportion to their molecular masses.
Many components of the translational machinery have been identified in C. elegans, including rRNAs (Albertson, 1984; Ellis et al., 1986; Files and Hirsh, 1981), ribosomal proteins (Jones and Candido, 1993; Zorio et al., 1994; Gonczy et al., 2000), 5S RNA (Nelson and Honda, 1985), tRNA (Schaller et al., 1991; Tranquilla et al., 1982; Khosla and Honda, 1989; Lee et al., 1990), and aminoacyl tRNA synthetases (Amaar and Baillie, 1993; Gabius et al., 1983; Gonczy et al., 2000).
Of the 56 initiation factor (eIF), elongation factor (eEF), and release factor (eRF) polypeptides that we and others have identified (Table 1), only ~10% have been isolated and characterized. In the following overview, information not explicitly cited is taken from WormBase, release WS138.
Table 1. C. elegans translational initiation, elongation, and termination factors
| Predicted translation factor | Transcript (splice variants) | CGC/ Other names | Predicted identity or function | Number of amino acids | Identity to C. briggsae/H. sapiens homologs (%) |
|---|---|---|---|---|---|
| Initiation factors | |||||
| eIF1 | T27F7.3b | -/PIG-B | Similar to SUI1 | 109 | 100/65 |
| eIF1A | H06H21.3 | – | Translation initiation factor 1A | 216 | 72/65 |
| eIF2A | E04D5.1 (a, b) | – | Contains similarity to translation initiation factor eIF2A | 618, 225 | 96/34 |
| eIF2α | Y37E3.10 | – | Translation initiation factor 2, α subunit | 342 | 88/47 |
| eIF2α | K04G2.1 | – | Translation initiation factor 2, α subunit | 250 | 82/58 |
| eIF2γ | Y39G10AR.8 | – | Translation initiation factor 2, γ subunit | 469 | 97/72 |
| eIF2B | C01G10.9 | – | Translation initiation factor related to eIF-2B,α/β/δ subunits | 366 | 84/43 |
| eIF2Bα | ZK1098.4 | – | eIF-2B,α subunit/GCN3 | 305 | 94/42 |
| eIF2Bβ | Y47H9C.7 | – | eIF-2B,β subunit | 340 | 99.7/25 |
| eIF2Bε | D2085.3 | – | eIF-2Bε subunit/ GCD6 | 666 | 99.8/23 |
| eIF2Bγ | C15F1.4 | PPP-1 | Translation initiation factor 2B, γ subunit/ pyrophosphorylase family | 404 | 79/29 |
| eIF2C-1 | T22B3.2 (a, b) | – | Translation initiation factor 2C and related proteins | 1032, 1035 | 99.9/36 |
| eIF2C-2 | ZK757.3 (a, b, c) | TAG-76 | Translation initiation factor 2C and related proteins | 1040, 1037, 55 | 99.9/42 |
| eIF2C-3 | R09A1.1 | – | Translation initiation factor 2C | 1121 | 98/27 |
| eIF3a | C27D11.1 | EGL-45/eif-3.A | Homologs to eIF3a | 1076 | 72/37 |
| eIF3a | F55H2.6 | CLU-1 | Involved in mitochondrial morphology/ distribution | 1247 | 80/40 |
| eIF3b | Y54E2A.11a | EIF-3.B | Translation initiation factor 3, subunit b | 725 | 89/34 |
| eIF3c | T23D8.4 | EIF-3.C | Translation initiation factor 3, subunit c | 898 | 89/39 |
| eIF3d | R08D7.3 | EIF-3.D | Translation initiation factor 3, subunit d | 570 | 94/45 |
| eIF3e | B0511.10 | EIF-3.E | Translation initiation factor 3, subunit e | 432 | 74/49 |
| eIF3f | D2013.7 | EIF-3.F | Translation initiation factor 3, subunit f | 294 | 61/35 |
| eIF3g | F22B5.2 | EIF-3.G | Translation initiation factor 3, subunit g | 256 | 90/31 |
| eIF3h | C41D11.2 | EIF-3.H | Translation initiation factor 3, subunit h | 365 | 97/39 |
| eIF3i | Y74C10AR.1 | EIF-3.I | Translation initiation factor 3, subunit i | 327 | 93/43 |
| eIF3j | Y40B1B.5 | – | Translation initiation factor eIF3, p35 subunit | 212 | 69/20 |
| eIF3k | T16G1.11 | EIF-3.K/pqn-69 | Eukaryotic initiation factor 3, p25 subunit | 240 | 89/35 |
| eIF3l | C17G10.9 (a.1, a.2, b) | – | RNA polymerase I-associated factor - PAF67 | 535-537 | 99/41 |
| eIF4A | F57B9.6 | INF-1 | Protein with high similarity to eukaryotic initiation factor 4A | 402 | 97/72 to eIF4A-2, 71 to eIF4A-1 |
| eIF4A | F57B9.3 | – | Translation initiation factor 4F, helicase subunit (eIF4A) and related helicases | 363 | 62/52 to eIF4A-1, 51 to eIF4A-2 |
| eIF4A-3 | Y65B4A.6 | – | ATP-dependent RNA helicase FAL1, involved in rRNA maturation | 399 | 93/82 to eIF4A-3, 66 to eIF4A-2, 61 to eIF4A-1 |
| eIF4A-3 | F33D11.10 | – | ATP-dependent RNA helicase FAL1, involved in rRNA maturation | 399 | 94/81 to eIF4A-3, 66 to eIF4A-2, 62 to eIF4A-1 |
| eIF4A-p56 | C07H6.5 | CGH-1 | Conserved germline helicase | 430 | 97/71 to DDX6 (p56); 40 to eIF4A-1 |
| eIF4A-47 | C26D10.2 (a, b) | HEL-1 | ATP-dependent RNA helicase | 425/268 | 97/78 to nuclear BAT1 (p47) |
| eIF4A-DDX47 | T26G10.1 | – | ATP-dependent RNA helicase | 489 | -/61 to DDX47 |
| eIF4A-DDX19 | T07D4.4 (a, b, c) | – | ATP-dependent RNA helicase | 1022, 638, 613 | 99.8/49 to DDX19 |
| eIF4A-Prp5 | F53H1.1 (a, b) | – | RNA helicase | 970, 747 | 99.9/46 to Prp5 |
| eIF4B | Y73B6BL.33 | f3 | Splicing factor hnRNP-F and related RNA-binding proteins | 610 | 62/31 |
| eIF4E-1 | F53A2.6 | IFE-1 | mRNA cap-binding protein | 212 | 95/40 |
| eIF4E-2 | R04A9.4 | IFE-2 | mRNA cap-binding protein | 228 | 98/40 |
| eIF4E-3 | B0348.6 (a, b, c) | IFE-3 | mRNA cap-binding protein | 248, 251, 250 | 99/47 |
| eIF4E-4 | C05D9.5 | IFE-4 | mRNA cap-binding protein | 212 | 95/30 to eIF4E and 48% to 4E-HP |
| eIF4E-5 | Y57A10A.30 (a, b) | IFE-5 | mRNA cap-binding protein | 240,201 | 80/40 |
| eIF4G | M110.4 (a, b) | IFG-1 | Translation initiation factor 4F, ribosome/ mRNA-bridging subunit | 1155, 1156 | 99/27 |
| eIF4H | T12D8.2 | – | mRNA cleavage and polyadenylation factor I complex, subunit RNA15 | 207 | 74/33 |
| eIF5 | C37C3.2 (a, b, c) | – | Translation initiation factor 5 | 436, 402, 413 | 99/48 |
| eIF5A | F54C9.1 | IFF-2 | Translation initiation factor 5 homolog | 161 | 99/61 |
| eIF5A | T05G5.10 | IFF-1 | Translation initiation factor 5 homolog | 161 | 99/57 |
| eIF5B | Y54F10BM.2 | – | Translation initiation factor 5B | 1173 | 82/48 |
| Elongation factors | |||||
| eEF1A | F31E3.5 | EFT-3 | Translation elongation factor 1α | 436 | 99.8/83 |
| eEF1A | R03G5.1 (a, b, c, d) | EFT-4 | Translation elongation factor 1α | 463, 77, 267, 429 | 99.8/84 |
| eEF1B | Y41E3.10 | – | Elongation factor 1 β/δ chain | 285 | 50.5/40 |
| eEF1B | F54H12.6 | – | Elongation factor 1 β/δ chain | 213 | 99.5/38 |
| eEF2 | F25H5.4 | EFT-2 | Homolog of translation elongation factor 2 | 852 | 89/76 |
| eEF2 | ZK328.2 | EFT-1 | Elongation factor 2 | 974 | 99/38 |
| Termination factors | |||||
| eRF1 | T05H4.6(a, b) | – | Peptide chain release factor 1 | 443, 559 | 99.8/80 |
| eRF3 | H19N07.1 | – | Peptide chain release factor 3 | 532 | 99.8/63 |
eIF2 factors. Both eIF2 and eIF2A catalyze the binding of Met-tRNAi to the 40S ribosomal subunit, the former requiring GTP and the latter, an AUG codon (Zoll et al., 2002). eIF2 is composed of three subunits, α, β, and γ. eIF2β has also been reported to bind mRNA and contributes, together with eIF2γ, to GTP and Met-tRNAi binding. Inactivation of either of these genes is lethal. Surprisingly, eIF2α deletion mutants are viable but have defects in growth and larval development. Yeast eIF2A is not an essential protein, but deletion of C. elegans eIF2A is embryonically lethal. eIF2B is a heteropentameric complex that is essential in yeast and mammalian cells. In C. elegans, inactivation of either the β, γ (Kuwabara and Shah, 1994), or ε subunits causes growth defects, larval arrest, or embryonic lethality.
eIF4 factors. Two splice-variants of eIF4G, encoded by ifg-1, have been identified in C. elegans (Long et al., 2002; Kamath et al., 2003). Although IFG-1 shows low identity to human eIF4G, its role as a translation factor was confirmed by its retention on m7GTP-Sepharose (Keiper and Rhoads, unpublished data) and by its presence in 48S initiation complexes (Dinkova et al., 2005). ifg-1 inactivation results in developmental arrest and embryonic lethality (Long et al., 2002).
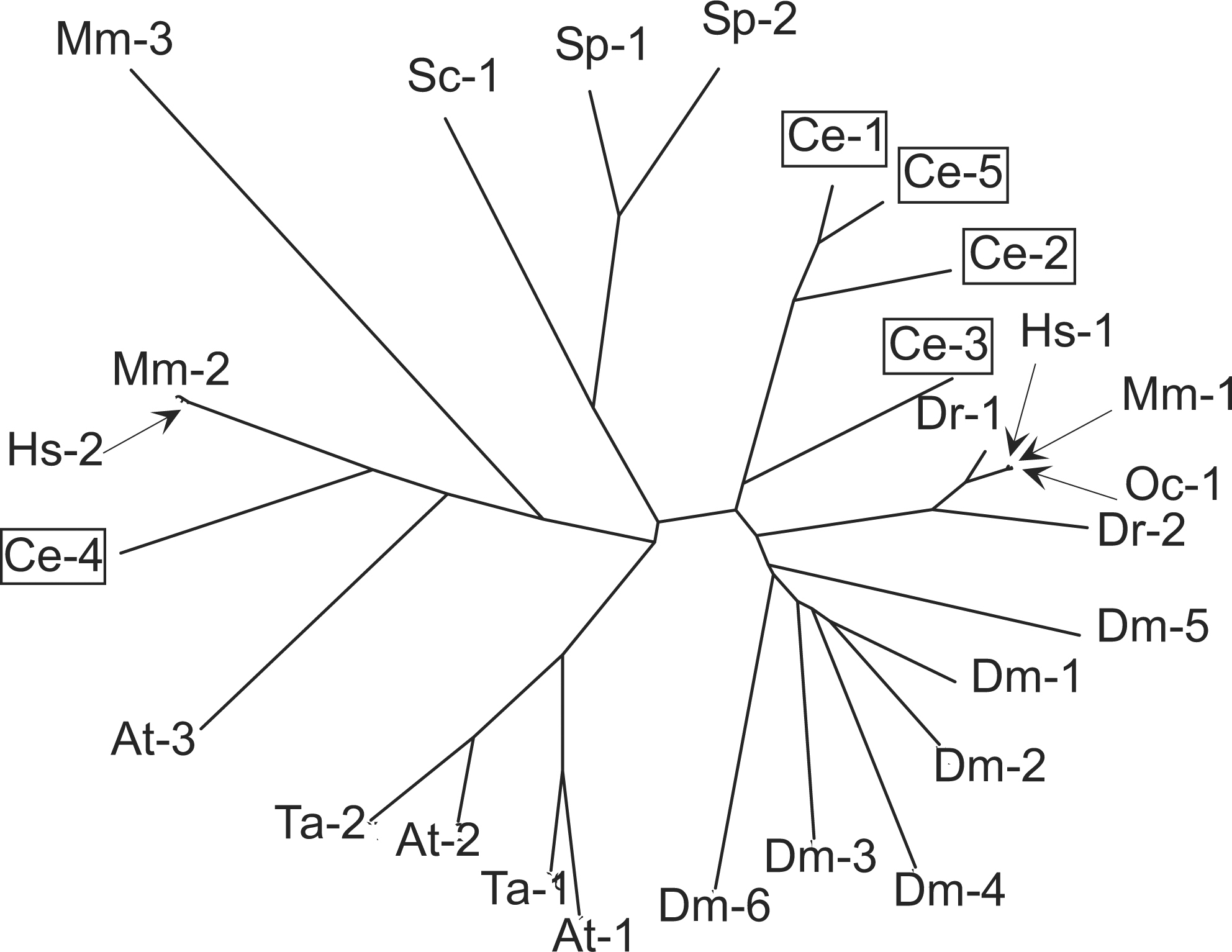
eIF4E is encoded by five genes in C. elegans, ife-1 through ife-5 (Jankowska-Anyszka et al., 1998; Keiper et al., 2000; Figure 3). The five proteins can be grouped into three classes based on amino acid sequence identity, cap-binding specificity, and knockout phenotype (Table 2). Some isoforms bind both 2,2,7-trimethylguanosine (TMG)-containing caps as well as 7-methylguanosine (MMG)-containing caps, whereas other isoforms bind only the latter. Cap selectivity is determined by the dimensions and flexibility of the cap-binding pocket (Miyoshi et al., 2002). IFE-1 is bound to P granules through an interaction with PGL-1 and is required for spermatogenesis (Amiri et al., 2001). In the case of IFE-4, inactivation of the gene produces an Egl phenotype, low brood size, and a defect in food sensation (Dinkova et al., 2005). The translational efficiency (polysomal distribution) of only ~1% of mRNAs is affected by ife-4 deletion, but diminished levels of the encoded proteins are consistent with the complex phenotype.
Table 2. Properties of five C. elegans isoforms of eIF4E
| Class | eIF4E isoform | Cap-binding specificity | RNAi phenotype | Tissue distribution | Knockout mutant phenotype | Postulated physiological role | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| A | IFE-3 | MMG | Lethal | Soma Germ line | Emb | General translation | ||||
| B | IFE-1 |
|
|
Germ line | Spe | Regulation of P granules mRNA translation | ||||
| IFE-2 |
|
Viable[a] | Soma | Unknown | Unknown | |||||
| IFE-5 |
|
Viable[a] | Unknown | Unknown | Unknown | |||||
| C | IFE-4 | MMG |
|
|
Egl | Specific mRNA translation during development | ||||
| References | Jankowska- Anyszka et al., 1998; Keiper et al., 2000; Miyoshi et al., 2002; Stachelska et al., 2002 | Keiper et al., 2000; Amiri et al., 2001; Dinkova et al., 2005 | Amiri et al., 2001; Dinkova et al., 2005 | Amiri et al., 2001; Dinkova et al., 2005 | Amiri et al., 2001; Dinkova et al., 2005; Trutschl et al., 2005 | |||||
|
aThe combination of RNAi for all three IFEs from Class B is lethal. |
||||||||||
|
Figure 3. Homologies among eIF4E isoforms. Amino acid sequence alignments were performed using Vector NTI 9.0.0 with a gap penalty of 3.0 and a gap extension penalty of 0.1. Multiple sequence alignments were used to calculate protein distance values and construct a phylogenetic tree with PHYLIP software on the Institut Pasteur server. eIF4E isoforms from C. elegans are indicated in boxes. NCBI accession numbers are indicated. A. thaliana: At-1, Y10548; At-2, Y10547; At-3, AF028809. C. elegans: Ce-1, NM_067350; Ce-2, NM_075693; Ce-3, NM_171920; Ce-4, AF214651; Ce-5, NM_064207. D. rerio: Dr-1, AF257519; Dr-2, AF176317. D. melanogaster: Dm-1, NM_168334; Dm-2, NM_139795; Dm-3, NM_139903; Dm-4, NM_166870; Dm-5, NM_143397; Dm-6, NM_139937. H. sapiens: Hs-1, M15353; Hs-2, AF047695. M. musculus: Mm-1, M61731; Mm-2, AF068116; Mm-3, AK005054 O.cuniculus: Oc-1, X61939. S. cerevisiae: Sc-1, M15436. S. pombe: Sp-1, X99444; Sp-2, AL031852, T. aestivum: Ta-1, Z12616; Ta-2, M95819.
Nine C. elegans gene products are homologous to eIF4A (Table 1; Roussell and Bennett, 1992; Gonczy et al., 2000). The products of F57B9.6 and F57B9.3 are the most homologous to mammalian eIF4A-1 and eIF4A-2. Others (Y65B4A.6 and F33D11.10) may be involved in splicing or nonsense-mediated decay, similar to mammalian eIF4A-3. Some may play a specific role in germline cells (C07H6.5) or in nuclear export (C26D10.2, T26G10.1, T07D4.4, and F53H1.1). Knockout of any eIF4A-like gene except T07D4.4 causes reproductive defects or embryonic lethality.
Other initiation factors and interacting proteins. eIF1, eIF3, and eIF5 form a multifactor complex in yeast that is required for assembly of the 43S initiation complex. C. elegans eIF5 and eIF1 are essential for growth and embryonic development. There are C. elegans homologs for all 12 eIF3 subunits present in higher eukaryotes (Asano et al., 1997), whereas yeast eIF3 has only five subunits. Loss of eIF3a (C27D11.1) produces an Egl phenotype (Desai and Horvitz, 1989). Both eIF3a and eIF3d are involved in meiotic divisions (Gonczy et al., 2000). Another eIF3a homolog (F55H2.6) is possibly involved in mitochondrial morphology and distribution. Inactivation of eIF5B results in larval arrest and sterility (Maeda et al., 2001). C. elegans contains three PABP homologs. PAB-2 (F18H3.3) has the highest similarity to human cytoplasmic PABP. PAB-1 (Y106G6H.2) is essential for gonad development, and PAB-2 is apparently important for somatic development (Ciosk et al., 2004). PAB-3 (C17E4.5) is more similar to human nuclear PABP and is probably not involved in translation.
Elongation and termination factors C. elegans has two identical eEF1A homologs that are products of different genes, eft-3 (Seydoux and Fire, 1994) and eft-4 (Kamath et al., 2003). EFT-3 is required for embryonic viability, fertility, and germline maintenance. There are two homologs of eEF1B, one of which (F54H12.6) is essential for viability while the other (Y41E3.10) is not. The two eEF2 homologs, EFT-1 (Ofulue and Candido, 1992) and EFT-2 (Ofulue and Candido, 1991; Nollen et al., 2004), are expressed during all stages and are encoded by essential genes. eRF1 has two splice variants whereas eRF3 has only one. Both genes are essential for embryonic and larval development, growth, and locomotion.
A distinctive feature of C. elegans is the presence of polycistronic pre-mRNA transcribed from operons (Blumenthal and Gleason, 2003). Translational control of individual mRNAs relies on cis-regulatory elements that include the cap, the 5'-terminal spliced leader (SL), and additional sequence elements in 5'- and 3'-untranslated regions (UTRs).
Spliced leaders. A SL is added to mature mRNA via trans-splicing in nematodes and some other metazoans (Bektesh et al., 1988; Evans et al., 1994; Stover and Steele, 2001). The SL consists of a conserved 22-nt sequence (Conrad et al., 1991; Blumenthal and Steward, 1997). In C. elegans, either SL1 or SL2 (or a variant such as SL3, SL4, etc.) is added to 70% of pre-mRNAs (Blumenthal and Gleason, 2003). SL1 is usually trans-spliced to mRNAs transcribed from the first cistron in an operon or to monocistronic mRNAs, whereas SL2 and its variants are trans-spliced to products of downstream cistrons.
Caps. Those mRNAs that undergo trans-splicing carry a TMG-cap and SL, whereas those that undergo only. cis-splicing carry an MMG-cap and no SL (Van Doren and Hirsh, 1990). The small nuclear RNA that donates the SL contains a TMG-cap (Zorio et al., 1994; Evans et al., 1997). MMG- and TMG-caps are recognized by some but not all eIF4E isoforms (Table 2).
5'UTR. Regulatory features of the 5'UTRs of mRNAs in general involve length, secondary structure, upstream open-reading frames, and specific sequences that interact with RNA binding proteins (Gingras et al., 1999). The trans-splicing processing of many C. elegans mRNAs results in a relatively short 5'UTR in which the SL is located near the AUG codon (Blumenthal and Steward, 1997). A few C. elegans mRNAs, e.g., gna-2, contain long 5'UTRs that harbor upstream open-reading frames (Lee and Schedl, 2004), which have a profound effect on translation efficiency in other organisims (Morris and Geballe, 2000).
Effects of 5'-terminal structures on translational efficiency. Mechanisms of regulation mediated by 5'-terminal structures remain largely unknown for C. elegans. Both MMG- and TMG-containing mRNAs are found on polysomes (Liou and Blumenthal, 1990). In Ascaris lumbricoides, mutations in the SL1 sequence alter efficiency of translation (Maroney et al., 1995). TMG-capped mRNAs are poorly translated in mammalian cell-free translational systems (Darzynkiewicz et al., 1988), but a TMG-cap and SL stimulate translation in an Ascaris suum translation system (Lall et al., 2004). Interestingly, an optimal distance from SL to AUG for translational efficiency can be demonstrated in this system. The translational synergism between cap and poly(A) is greater for TMG than MMG. The well described translational regulator GLD-1 also interacts with 5'UTR sequences (Lee and Schedl, 2004).
3'UTR. Translational regulatory elements at the 3'UTR play important roles in C. elegans mRNA expression (see WormBook chapters on Translational control of maternal RNAs and RNA-binding proteins). The elements first described as DREs (direct repeat elements; Goodwin et al., 1993) and later re-named TGEs (tra GLI elements; Jan et al., 1997) are found within the 3'UTR of tra-2 mRNA and negatively regulate its expression in germ line and somatic cells. The trans-acting factor for TGEs was identified as GLD-1 (Jan et al., 1999). A 5-nucleotide sequence element in the 3'UTR of fem-3 mRNA, cuUCUUGu, also exerts translational regulation (Anderson and Kimble, 1997). Another kind of 3'UTR element that represses translation is the lin-4 complementary element (LCE) found in lin-14 and lin-28 mRNAs (Wightman et al., 1993; Seggerson et al., 2002). This element is bound by the microRNA lin-4 (Moss et al., 1997; see C. elegans microRNAs). Mutations in these 3'UTR elements disrupt the translational regulation of their mRNAs (see WormBook chapters referenced above).
poly(A) The poly(A) tract is added by a poly(A) polymerase after specific cleavage during mRNA splicing. The nuclear polyadenylation signal consists of an AAUAAA sequence 20-30 nt upstream of the cleavage site (Blumenthal, 1995; Hajarnavis et al., 2004). In C. elegans, GLD-2 is a putative catalytic subunit of cytoplasmic poly(A) polymerase that is likely recruited to mRNAs by interaction with RNA binding proteins such as GLD-3 (Wang et al., 2002). Cytoplasmic polyadenylation also requires a cytoplasmic polyadenylation element (CPE), located upstream of the polyadenylation site, which is recognized by specific proteins (CPEBs).
Effects of 3'-terminal structures on tranlational efficiency. Although there are many instances in which specific structures in the 3'UTR have been shown to affect translational efficiency in C. elegans and other organisms (Kuersten and Goodwin, 2003), the molecular interactions responsible for these effects are only partially understood. In Xenopus oocytes, CPEB binds and sequesters eIF4E through an intermediary protein, Maskin (Mendez and Richter, 2001). In Drosophila embryos, there is a similar interaction between the 3'UTR-binding factor Smaug and eIF4E, mediated by another protein, Cup (Nelson et al., 2004). However, in C. elegans the translational component(s) involved in GLD-1-mediated regulation remain unknown (Jan et al., 1999; Marin and Evans, 2003; Lee and Schedl, 2004). The other 3'-terminal element, the poly(A) tract, increases the rate of translational initiation in yeast and plants due to the binding of PABP to a specific site near the N-terminus of eIF4G (Tarun and Sachs, 1996; Le et al., 1997; see Figure 2). Poly(A) stabilizes the PABP•eIF4G•eIF4E complex, which in turn leads to enhanced translational re-initiation (Wakiyama et al., 2000). As discussed below, there are several regulatory mechanisms in C. elegans that involve changing the poly(A) length.
Many of the translational mechanisms that have been well described and characterized in other organisms (Sonenberg et al., 2000) have not yet been demonstrated in C. elegans. These can be divided into two broad classes, modification of translation factors and modification of mRNA structure. However, the existence of homologous translation factors (Table 1) and signal transduction components (Table 3) suggests that previously discovered mechanisms operate in C. elegans as well.
Table 3. Components of signal transduction pathways implicated in translational control
| Predicted signal transduction component | Transcript (splice variants) | CGC/ Other names | Predicted identity or function (WormBase, release WS138) |
|---|---|---|---|
| P70S6K | Y47D3A.4 | CKU-70 | DNA-binding subunit of a DNA-dependent protein kinase |
| TAP42 | Y71H2B.3 | Protein phosphatase 2A-associated protein | |
| TIP41 | ZK688.9 | Uncharacterized conserved protein | |
| SIT4.1 | Y75B8A.30 | PPH-4.1 | Serine/threonine-specific protein phosphatase involved in glycogen accumulation |
| SIT4.2 | Y49E10.3a | PPH-4.2 | Serine/threonine-specific protein phosphatase involved in glycogen accumulation |
| Akt | C12D8.10 (a, b, c) | AKT-1 | Ortholog of serine/threonine kinase Akt/PKB |
| PERK | F46C3.1 | PEK-1 | Human PERK kinase homolog |
| PEK | Y38E10A.8 | eIF2α kinase PEK/EIF2AK3 | |
| MKK7/JNKK2 | K08A8.1 | MEK-1/ KIN-17 | MKK7/JNKK2 |
| MKK7/JNKK2 | F35C8.3 | JKK-1 | JNK Kinase |
| MKK4 | F42G10.2 | MKK-4 | MKK (MAP kinase kinase) homolog |
| MAPK7/ERK5 | W06B3.2 | SMA-5 | MAPK7/ERK5 |
| MAPK7 | C04G6.1 | MAP-2 | Mitogen-activated protein kinase 7, isoform 1 |
| MAPK7 | F09C12.2 | Mitogen-activated protein kinase | |
| MAP2K | Y54E10BL.6 | MEK-2/ LET-537/ GLV-1 | MAP kinase kinase or ERK kinase |
| ERN1/IRE1 | C41C4.4 | IRE-1 | IRE1 kinase related |
| TOR | B0261.2 | LET-363 | Ortholog of S. cerevisiae Tor1p and Tor2p and human FRAP1 |
| PI3K | C46B6.6 | SMG-1/ MAB-1 | PI-3-kinase-related kinase SMG-1 isoform 2 |
| PI3K like | T06E4.3 | ATL-1 | ATM (ataxia telangectasia mutated)-like |
| PI4K | ZC8.6 | Phosphatidylinositol 4-kinase | |
| PI4K | C56A3.8 | Phosphatidylinositol 4-kinase | |
| Raptor | C10C5.6 (a, b) | DAF-15 | Abnormal dauer formation |
Covalent modification of a translation factor. The unfolded protein response (UPR) is a transcriptional and translational signaling pathway activated by the accumulation of unfolded proteins in the endoplasmic reticulum (ER; Zhang and Kaufman, 2004). This involves activation of an eIF2α kinase, PEK, causing global inhibition of translation initiation and allowing time to remedy the folding problem. C. elegans PEK-1 was expressed in yeast and found to inhibit growth by hyperphosphorylation of eIF2α and inhibition of eIF2B (Sood et al., 2000). UPR gene transcription and survival upon ER stress also requires ire-1-mediated splicing of xbp-1 mRNA (Shen et al., 2001). ire-1/xbp-1 acts with pek-1 in complementary pathways that are essential for worm development, survival, and ER homeostasis. Furthermore, pek-1 mutants have shortened life spans (Harding et al., 2003).
Sequestration of a translation factor. TOR is a highly conserved protein kinase that controls cell growth and division in eukaryotes. In mammals, mTOR regulates translation by phosphorylation of p70 S6 kinase (S6K) and the eIF4E-binding protein 4E-BP1 (Gingras et al., 2001). The latter event releases eIF4E from a sequestered form, making it available to bind eIF4G (Figure 2). Raptor is a mTOR-binding protein that is necessary for the mTOR-catalyzed phosphorylation of 4E-BP1 and S6K (Hara et al., 2002). In yeast, TOR also regulates translation through eIF4E by a mechanism involving the phosphatases Sit4 and PP2A and the phosphatase-binding protein Tap42 (Jiang and Broach, 1999). Tip41 negatively regulates TOR by binding and inhibiting Tap42 (Jacinto et al., 2001).
In C. elegans, cTOR (let-363) deficiency causes developmental arrest and intestinal atrophy (Long et al., 2002). The phenotype resembles that of RNAi knockdown of eIF4G, eIF2α, and eIF2β, but not S6K, Tip41, or Tap42. RNAi of Raptor (daf-15) yields an array of phenotypes resembling those of cTOR knockout. Deficiency of both let-363 (Vellai et al., 2003) and daf-15 (Jia et al., 2004) extends adult lifespan, while mutations in either gene result in dauer-like larval arrest (Jia et al., 2004). To date, a C. elegans 4E-BP1 homolog has not been identified. The protein phas-1 (WP:CE30964; Agostoni et al., 2002; another name for 4E-BP1 is PHAS-I) is unrelated to mammalian and Drosophila 4E-BP1.
The presence of IFE-1 in P granules (Amiri et al., 2001) and its absence from initiation complexes (Dinkova et al., 2005) may represent another regulatory mechanism involving sequestration of a translation factor. A number of mRNAs have been found in P granules, some of which have been shown to be translationally controlled (Schisa et al., 2001). P granules also contain four GLH RNA helicases similar to eIF4A (Gruidl et al., 1996; Kuznicki et al., 2000). In Drosophila, VASA is a germline-specific ATP-dependent RNA helicase, homologous to the GLH proteins, that is required for translation of at least two mRNAs, gurken and nanos, through interaction with eIF5B (Styhler et al., 1998; Johnstone and Lasko, 2004). Also, eIF5A is required for PGL-1 localization (Hanazawa et al., 2004). The presence of all these translational components in P granules may signify regulation of germline-specific mRNA translation.
The translation of some C. elegans mRNAs is regulated by 3'UTR-binding proteins that alter poly(A) length (Kuersten and Goodwin, 2003). There are four CPEB homologs, CBP-1, CPB-2, CPB-3, and FOG-1 (Luitjens et al., 2000). CPB-1 is essential for progression through meiosis and is present in germ cells just before spermatogenesis. CPB-1 physically interacts with FBF, another RNA-binding protein and 3' UTR regulator. Similarities between FOG-1 and the CPEB proteins of Xenopus and Drosophila suggest that FOG-1 controls germ cell fates by regulating the translation of specific messenger RNAs.
A specific mRNA that undergoes regulated changes in poly(A) length is tra-2 mRNA, whose translation must be repressed for male development. A possible mechanism of action is suggested by the observation that TGEs control the length of the poly(A) tract in a Xenopus system (Thompson et al., 2000). Similarly, a 5-nt element in the 3'UTR of fem-3 mRNA controls the length of the poly(A) tract (Ahringer and Kimble, 1991). fem-3 must be downregulated to allow the switch from spermatogenesis to oogenesis. FBF and Nanos-3 interact with each other and repress translation of fem-3 mRNA (Kraemer et al., 1999). GLD-3 also physically interacts with FBF to interfere with FBF binding to the 3'UTR of fem-3 mRNA (Eckmann et al., 2002). GLD-3 promotes the transition from mitosis to meiosis together with the putative GLD-2 poly(A) polymerase. FBF also binds specifically to elements in the 3'UTR of gld-3S mRNA and regulates gld-3 expression (Eckmann et al., 2004).
The translation of several other mRNAs is known to be regulated through the 3'UTR, but this cannot yet be explained within the framework of known translational control mechanisms. A 34-nucleotide region of the 3'UTR of glp-1 mRNA contains two regulatory elements, one that represses translation in germ cells and posterior cells of the early embryo, and one that inhibits repressor activity to promote translation in the embryo (Marin and Evans, 2003). GLD-1 binds to this repressor element. MEX-3 is also expressed in anterior blastomere cells and is required for repression of pal-1 mRNA translation (Hunter and Kenyon, 1996; Draper et al., 1996).
A different type of 3'UTR regulation is seen with the heterochronic genes, which are temporally controlled to specify the timing of developmental events. lin-14 and lin-28 encode proteins that are required for temporal execution of cell lineages during larval development. lin-14, lin-28, lin-42, and daf-12 mRNAs contain a conserved element in the 3'UTR, the loss of which causes a gain of function (Seggerson et al., 2002). let-7 encodes a temporally regulated 21-nucleotide RNA that is complementary to elements in heterochonic mRNA 3'UTRs and regulates their translation (Reinhart et al., 2000). A second regulatory RNA, lin-4, negatively regulates lin-14 and lin-28 through RNA-RNA interactions with their 3'UTR (Moss et al., 1997). lin-28 is repressed during normal development by a mechanism that acts on its mRNA after translation initiation (Seggerson et al., 2002).
As illustrated throughout this chapter, several key developmental pathways in C. elegans are predominantly governed by translational mechanism. However, translation per se has received little attention. Even though one can find C. elegans homologs for most of the RNAs and proteins that make up the translational machinery (Table 1), few of these have been isolated, functionally identified, or studied at the biochemical level. This deficiency obscures the molecular mechanisms responsible for well characterized physiological processes. Another shortcoming is the absence of a cell-free translation system, although such systems have been developed in two other nematodes, Ascaris lumbricoides (Maroney et al., 1995) and Ascaris suum (Lall et al., 2004). The most interesting unexplored questions may involve changes in the translation of individual mRNAs in various physiological conditions or after genetic manipulations. Given the highly developed state of C. elegans genomics, it can be expected that future application of computational tools, including data visualization (Trutschl et al., 2005), will help detect new instances of translational control.
This work was supported by Grant GM20818 from the National Institutes of Health. We thank Brett Keiper for sharing preliminary data and for helpful discussions, Aaron Lefebvre for protein sequence alignments and construction of Figure 3, and Lisa Whittington for editorial assistance.
Agostoni, E., Gobessi, S., Petrini, E., Monte, M., and Schneider, C. (2002). Cloning and characterization of the C. elegans gas1 homolog: phas-1. Biochim. Biophys. Acta 1574, 1–9. Abstract
Ahringer, J., and Kimble, J. (1991). Control of the sperm-oocyte switch in Caenorhabditis elegans hermaphrodites by the fem-3 3' untranslated region. Nature 349, 346–348. Abstract Article
Albertson, D. (1984). Localization of the ribosomal genes in Caenorhabditis elegans chromosomes by in situ hybridization using biotin-labeled probes. EMBO J. 3, 1227–1234. Abstract
Amaar, Y., and Baillie, D. (1993).Cloning and characterization of the C. elegans histidyl-tRNA synthetase gene [published erratum appears in Nucleic Acids Res. 1993 Dec 25;21(25):6050-1]. Nucl. Acids Res. 21, 4344–4347. Abstract
Amiri, A., Keiper, B.D.,Kawasaki, I., Fan, Y., Kohara, Y., Rhoads, R.E., and Strome, S. (2001). An isoform of eIF4E is a component of germ granules and is required for spermatogenesis in C. elegans.Development 128, 3899–3912. Abstract
Anderson, P., and Kimble, J. (1997). mRNA and translation. In C. Elegans II, D.L. Riddle, T. Blumenthal, B.J. Meyer,and J.R. Priess,eds. (Cold Spring Harbor: Cold Spring Harbor Laboratory Press), pp. 185–208.
Asano, K., Clayton, J., Shalev, A., and Hinnebusch, A.G. (2000). A multifactor complex of eukaryotic initiation factors, eIF1, eIF2, eIF3, eIF5, and initiator tRNA(Met) is an important translation initiation intermediate in vivo. Genes Dev. 14, 2534–2546. Abstract Article
Asano, K., Vornlocher, H.-P., Richter-Cook, N.J., Merrick, W.C., Hinnebusch, A.G., and Hershey, J.W.B. (1997). Structure of cDNAs encoding human eukaryotic initiation factor 3 subunits. J. Biol. Chem. 272, 27042–27052. Abstract Article
Bektesh, S., Van Doren, K., and Hirsh, D. (1988). Presence of the Caenorhabditis elegans spliced leader on different mRNAs and in different genera of nematodes. Genes Dev. 2, 1277–1283. Abstract
Blumenthal, T. (1995). Trans-splicing and polycistronic transcription in Caenorhabditis elegans. Trends Genet. 11, 132–136. Abstract Article
Blumenthal, T., and Gleason, K. (2003). Caenorhabditis elegans operons: form and function. Nat. Rev. Genet. 4, 112–120. Abstract Article
Blumenthal, T., and Steward, K. (1997). RNA processing and gene structure. In: C. Elegans II, D.L. Riddle, T. Blumenthal, B.J. Meyer, and J.R. Priess, eds. (Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press), pp. 117–145.
Ciosk, R., DePalma, M., and Priess, J.R. (2004). ATX-2, the C. elegans ortholog of ataxin 2, functions in translational regulation in the germline. Development 131, 4831–4841. Abstract Article
Clark, B.F.C., Grunberg-Manago, M., Gupta, N.K., Hershey, J.W.B., Hinnebusch, A.G., Jackson, R.J., Maitra, U., Mathews, M.B., Merrick, W.C., Rhoads, R.E., Sonenberg, N., Spremulli, L.L., Trachsel, H., and Voorma, H.O. (1996). Prokaryotic and eukaryotic translation factors. Biochimie 78, 1119–1122. Abstract Article
Conrad, R., Thomas, J., Spieth, J., and Blumenthal, T. (1991). Insertion of part of an intron into the 5' untranslated region of a Caenorhabditis elegans gene converts it into a trans-spliced gene. Mol. Cell. Biol. 11, 1921–1926. Abstract
Darzynkiewicz, E., Stepinski, J., Ekiel, I., Jin, Y., Haber, D., Sijuwade, T., and Tahara, S.M. (1988). β-globin mRNAs capped with m7G, m2,2G, or m2,2,7G differ in intrinsic translation efficiency. Nucl. Acids Res. 16, 8953–8962. Abstract
Desai, C., and Horvitz, H.R. (1989). Caenorhabditis elegans mutants defective in the functioning of the motor neurons responsible for egg laying. Genetics 121, 703–721. Abstract
Dinkova, T.D., Keiper, B.D., Korneeva, N.L., Aamodt, E.J., and Rhoads, R.E. (2005). Translation of a small subset of Caenorhabditis elegans mRNAs is dependent on a specific eIF4E isoform. Mol. Cell Biol. 25, 100–113. Abstract Article
Draper, B.W., Mello, C.C., Bowerman, B., Hardin, J., and Priess, J.R. (1996). MEX-3 is a KH domain protein that regulates blastomere identity in early C. elegans embryos. Cell 87, 205–216. Abstract Article
Eckmann, C., Kraemer, B., Wickens, M., and Kimble, J. (2002). GLD-3, a bicaudal-C homolog that inhibits FBF to control germline sex determination in C. elegans. Dev. Cell 3, 697–710. Abstract Article
Eckmann, C.R., Crittenden, S.L., Suh, N., and Kimble, J. (2004). GLD-3 and control of the mitosis/meiosis decision in the germline of Caenorhabditis elegans. Genetics 168, 147–160. Abstract Article
Ellis, R., Sulston, J., and Coulson, A. (1986). The rDNA of C. elegans: sequence and structure. Nucl. Acids Res. 14, 2345–2364. Abstract
Evans, D., Zorio, D., MacMorris, M., Winter, C.E., Lea, K., and Blumenthal, T. (1997). Operons and SL2 trans-splicing exist in nematodes outside the genus Caenorhabditis. Proc. Natl. Acad. Sci. USA 94, 9751–9756. Abstract Article
Evans, T., Crittenden, S., Kodoyianni, V., and Kimble, J. (1994). Translational control of maternal glp-1 mRNA establishes an asymmetry in the C. elegans embryo. Cell 77, 183–194. Abstract Article
Files, J., and Hirsh, D. (1981). Ribosomal DNA of Caenorhabditis elegans. J. Mol. Biol. 149, 223–240. Abstract Article
Gabius, H., Graupner, G., and Cramer, F. (1983). Activity patterns of aminoacyl-tRNA synthetases, tRNA methylases, arginyltransferase and tubulin: tyrosine ligase during development and ageing of Caenorhabditis elegans. Eur. J. Biochem. 131, 231–234. Abstract Article
Gingras, A.-C., Raught, B., and Sonenberg, N. (1999). eIF4 initiation factors: effectors of mRNA recruitment to ribosomes and regulators of translation. Ann. Rev. Biochem. 68, 913–963. Abstract Article
Gingras, A.-C., Raught, B., and Sonenberg, N. (2001). Regulation of translation initiation by FRAP/mTOR. Genes Dev. 15, 807–826. Abstract Article
Gonczy, P., Echeverri, C., Oegema, K., Coulson, A., Jones, S., Copley, R., Duperon., J., Oegema, J., Brehm., M.,Cassin., E., Hannak, E., Kirkham, M., Pichler, S., Flohrs, K., Goessen, A., Leidel, S., Alleaume, A.M., Martin, C., Ozlu, N., Bork, P., and Hyman, A.A. (2000). Functional genomic analysis of cell division in C. elegans using RNAi of genes on chromosome III. Nature 408, 331–336. Abstract Article
Goodwin, E., Okkema, P., Evans, T., and Kimble, J. (1993). Translational regulation of tra-2 by its 3' untranslated region controls sexual identity in C. elegans. Cell 75, 329–339. Abstract Article
Gruidl, M.E., Smith, P.A., Kuznicki, K.A., McCrone, J.S., Kirchner, J., Roussell, D.L., Strome, S., and Bennett, K.L. (1996). Multiple potential germ-line helicases are components of the germ-line-specific P granules of Caenorhabditis elegans. Proc. Natl. Acad. Sci. USA 93, 13837–13842. Abstract Article
Hajarnavis, A., Korf, I., and Durbin, R. (2004). A probabilistic model of 3' end formation in Caenorhabditis elegans. Nucl. Acids Res. 32, 3392–3399. Abstract Article
Hanazawa, M., Kawasaki, I., Kunitomo, H., Gengyo-Ando, K., Bennett, K., Mitani, S., and Iino, Y. (2004). The Caenorhabditis elegans eukaryotic initiation factor 5A homologue, IFF-1, is required for germ cell proliferation, gametogenesis and localization of the P-granule component PGL-1. Mech. Dev. 121, 213–224. Abstract Article
Hara, K., Maruki, Y., Long, X., Yoshino, K., Oshiro, N., Hidayat, S., Tokunaga, C., Avruch, J., and Yonezawa, K. (2002). Raptor, a binding partner of target of rapamycin (TOR), mediates TOR action. Cell 110, 177–189. Abstract Article
Harding, H., Zhang, Y., Zeng, H., Novoa, I., Lu, P., Calfon, M., Sadri, N., Yun, C., Popko, B., Paules, R., et al. (2003). An integrated stress response regulates amino acid metabolism and resistance to oxidative stress. Mol. Cell 11, 619–633. Abstract Article
Hershey, J.W.B., and Merrick, W.C. (2000). Pathway and mechanism of initiation of protein synthesis. In: Translational Control of Gene Expression, N. Sonenberg, J.W.B. Hershey, and M.B. Mathews, eds. (Cold Spring Harbor, New York: Cold Spring Harbor Laboratory Press), pp. 33–88.
Hinnebusch, A.G. (1997). Translational regulation of yeast GCN4: a window on factors that control initiator-tRNA binding to the ribosome. J. Biol. Chem. 272, 21661–21664. Abstract Article
Hinnebusch, A.G., and Fink, G.R. (1983). Positive regulation in the general amino acid control of Saccharomyces cerevisiae. Proc. Natl. Acad. Sci. USA 80, 5374–5378. Abstract
Hunter, C., and Kenyon, C. (1996). Spatial and temporal controls target pal-1 blastomere-specification activity to a single blastomere lineage in C. elegans embryos. Cell 87, 217–226. Abstract Article
Jacinto, E., Guo, B., Arndt, K., Schmelzle, T., and Hall, M. (2001). TIP41 interacts with TAP42 and negatively regulates the TOR signaling pathway. Mol. Cell 8, 1017–1026. Abstract Article
Jan, E., Motzny, C.K., Graves, L.E., and Goodwin, E.B. (1999). The STAR protein, GLD-1, is a translational regulator of sexual identity in Caenorhabditis elegans. EMBO J. 18, 258–269. Abstract Article
Jan, E., Yoon, J.W., Walterhouse, D., Iannaccone, P., and Goodwin, E.B. (1997). Conservation of the C. elegans tra-2 3'UTR translational control. EMBO J. 16, 6301–6313. Abstract Article
Jankowska-Anyszka, M., Lamphear, B.J., Aamodt, E.J., Harrington, T., Darzynkiewicz, E., Stolarski, R., and Rhoads, R.E. (1998). Multiple isoforms of eukaryotic protein synthesis initiation factor 4E in C. elegans can distinguish between mono- and trimethylated mRNA cap structures. J. Biol. Chem. 273, 10538–10542. Abstract Article
Jia, K., Chen, D., and Riddle, D.L. (2004). The TOR pathway interacts with the insulin signaling pathway to regulate C. elegans larval development, metabolism and life span. Development 131, 3897–3906. Abstract Article
Jiang, Y., and Broach, J.R. (1999). Tor proteins and protein phosphatase 2A reciprocally regulate Tap42 in controlling cell growth in yeast. EMBO J. 18, 2782–2792. Abstract Article
Johnstone, O., and Lasko, P. (2004). Interaction with eIF5B is essential for Vasa function during development. Development 131, 4167–4178. Abstract Article
Jones, D., and Candido, E. (1993). Novel ubiquitin-like ribosomal protein fusion genes from the nematodes Caenorhabditis elegans and Caenorhabditis briggsae. J. Biol. Chem. 268, 19545–19551. Abstract
Kamath, R., Fraser, A., Dong, Y., Poulin, G., Durbin, R., Gotta, M., Kanapin, A., Le Bot, N., Moreno, S., Sohrmann, M., et al. (2003). Systematic functional analysis of the Caenorhabditis elegans genome using RNAi. Nature 421, 231–237. Abstract Article
Keiper, B.D., Lamphear, B.J., Deshpande, A.M., Jankowska-Anyszka, M., Aamodt, E.J., Blumenthal, T., and Rhoads, R.E. (2000). Functional characterization of five eIF4E isoforms in Caenorhabditis elegans. J. Biol. Chem. 275, 10590–10596. Abstract Article
Khosla, M., and Honda, B. (1989). Initiator tRNAMet genes from the nematode Caenorhabditis elegans. Gene 76, 321–330. Abstract Article
Kraemer, B., Crittenden, S., Gallegos, M., Moulder, G., Barstead, R., Kimble, J., and Wickens, M. (1999). NANOS-3 and FBF proteins physically interact to control the sperm-oocyte switch in Caenorhabditis elegans. Curr. Biol. 9, 1009–1018. Abstract Article
Kuersten, S., and Goodwin, E. (2003). The power of the 3' UTR: translational control and development. Nat. Rev. Genet. 4, 626–637. Abstract Article
Kuwabara, P., and Shah, S. (1994). Cloning by synteny: identifying C. briggsae homologues of C. elegans genes. Nucl. Acids Res. 22, 4414–4418. Abstract
Kuznicki, K., Smith, P., Leung-Chiu, W., Estevez, A., Scott, H., and Bennett, K. (2000). Combinatorial RNA interference indicates GLH-4 can compensate for GLH-1; these two P granule components are critical for fertility in C. elegans. Development 127, 2907–2916. Abstract
Lall, S., Friedmann, C.C., Jankowska-Anyszka, M., Stepinksi, J., Darzynkiewicz, E., and Davis, R.E. (2004). Contribution of trans-splicing, 5'-leader length, cap-poly(A) synergism, and initiation factors to nematode translation in an Ascaris suum embryo cell-free system. J. Biol. Chem. 279, 45573–45585. Abstract Article
Le, H., Tanguay, R.L., Balasta, M.L., Wei, C.C., Browning, K., Metz, A.M., Goss, D.J., and Gallie, D.R. (1997). Translation initiation factors eIF-iso4G and eIF-4B interact with the poly(A)-binding protein and increase its RNA binding activity. J. Biol. Chem. 272, 16247–16255. Abstract Article
Lee, B.J., Rajagopalan, M., Kim, Y.S., You, K.H., Jacobson, K.B., and Hatfield, D. (1990). Selenocysteine tRNA[Ser]Sec gene is ubiquitous within the animal kingdom. Mol. Cell Biol. 10, 1940–1949. Abstract
Lee, M.-H., and Schedl, T. (2004). Translation repression by GLD-1 protects its mRNA targets from nonsense-mediated mRNA decay in C. elegans. Genes Dev. 18, 1047–1059. Abstract Article
Liou, R.F., and Blumenthal, T. (1990). Trans-spliced Caenorhabditis elegans messenger RNAs retain trimethylguanosine caps. Mol. Cell Biol. 10, 1764–1768. Abstract
Long, X., Spycher, C., Han, Z., Rose, A., Muller, F., and Avruch, J. (2002). TOR deficiency in C. elegans causes developmental arrest and intestinal atrophy by inhibition of mRNA translation. Curr. Biol. 12, 1448–1461. Abstract Article
Luitjens, C., Gallegos, M., Kraemer, B., Kimble, J., and Wickens, M. (2000). CPEB proteins control two key steps in spermatogenesis in C. elegans. Genes Dev. 14, 2596–2609. Abstract Article
Maeda, I., Kohara, Y., Yamamoto, M., and Sugimoto, A. (2001). Large-scale analysis of gene function in Caenorhabditis elegans by high-throughput RNAi. Curr. Biol. 11, 171–176. Abstract Article
Marin, V.A., and Evans, T.C. (2003). Translational repression of a C. elegans Notch mRNA by the STAR/KH domain protein GLD-1. Development 130, 2623–2632. Abstract Article
Maroney, P.A., Denker, J.A., Darzynkiewicz, E., Laneve, R., and Nilsen, T.W. (1995). Most mRNAs in the nematode Ascaris lumbricoides are trans-spliced: a role for spliced leader addition in translational efficiency. RNA 1, 714–723. Abstract
Mendez, R., and Richter, J.D. (2001). Translational control by CPEB: a means to the end. Mol. Cell Biol. 2, 521–529. Abstract Article
Miyoshi, H., Dwyer, D.S., Keiper, B.D., Jankowska-Anyszka, M., Darzynkiewicz, E., and Rhoads, R.E. (2002). Discrimination between mono- and trimethylated cap structures by two isoforms of Caenorhabditis elegans eIF4E. EMBO J. 21, 1–11. Abstract Article
Morris, D.R., and Geballe, A.P. (2000). Upstream open reading frames as regulators of mRNA translation. Mol. Cell Biol. 20, 8635–8642. Abstract Article
Moss, E., Lee, R., and Ambros, V. (1997). The cold shock domain protein LIN-28 controls developmental timing in C. elegans and is regulated by the lin-4 RNA. Cell 88, 637–646. Abstract Article
Nelson, D., and Honda, B. (1985). Genes coding for 5S ribosomal RNA of the nematode Caenorhabditis elegans. Gene 38, 245–251. Abstract Article
Nelson, M.R., Leidal, A.M., and Smibert, C.A. (2004). Drosophila Cup is an eIF4E-binding protein that functions in Smaug-mediated translational repression. EMBO J. 23, 150–159. Abstract Article
Nollen, E.A.A., Garcia, S.M., van Haaften, G., Kim, S., Chavez, A., Morimoto, R.I., and Plasterk, R.H.A. (2004). From the cover: genome-wide RNA interference screen identifies previously undescribed regulators of polyglutamine aggregation. Proc. Natl. Acad. Sci. USA 101, 6403–6408. Abstract Article
Ofulue, E., and Candido, E. (1991). Molecular cloning and characterization of the Caenorhabditis elegans elongation factor 2 gene (eft-2). DNA Cell Biol. 10, 603–611. Abstract
Ofulue, E., and Candido, E. (1992). Isolation and characterization of eft-1, an elongation factor 2-like gene on chromosome III of Caenorhabditis elegans. DNA Cell Biol. 11, 71–82. Abstract
Phan, L., Zhang, X., Asano, K., Anderson, J., Vornlocher, H.-P., Greenberg, J.R., Qin, J., and Hinnebusch, A.G. (1998). Identification of a translation initiation factor 3 (eIF3) core complex, conserved in yeast and mammals, that interacts with eIF5. Mol. Cell Biol. 18, 4935–4946. Abstract
Reinhart, B., Slack, F., Basson, M., Pasquinelli, A., Bettinger, J., Rougvie, A., Horvitz, H., and Ruvkun, G. (2000). The 21-nucleotide let-7 RNA regulates developmental timing in Caenorhabditis elegans. Nature 403, 901–906. Abstract Article
Roussell, D., and Bennett, K. (1992). Caenorhabditis cDNA encodes an eIF-4A-like protein. Nucl. Acids Res. 20, 3783. Abstract
Schaller, D., Wittmann, C., Linning, R., Spicher, A., Muller, F., and Tobler, H. (1991). Cloning and expression in vitro of a gene encoding tRNAArgACG from the nematode Caenorhabditis elegans. Gene 97, 273–276. Abstract Article
Schisa, J., Pitt, J., and Priess, J. (2001). Analysis of RNA associated with P granules in germ cells of C. elegans adults. Development 128, 1287–1298. Abstract
Seggerson, K., Tang, L., and Moss, E. (2002). Two genetic circuits repress the Caenorhabditis elegans heterochronic gene lin-28 after translation initiation. Dev. Biol. 243, 215–225. Abstract Article
Seydoux, G., and Fire, A. (1994). Soma-germline asymmetry in the distributions of embryonic RNAs in Caenorhabditis elegans. Development 120, 2823–2834. Abstract
Shen, X., Ellis, R., Lee, K., Liu, C., Yang, K., Solomon, A., Yoshida, H., Morimoto, R., Kurnit, D., Mori, K., and Kaufman, R. (2001). Complementary signaling pathways regulate the unfolded protein response and are required for C. elegans development. Cell 107, 893–903. Abstract Article
Sherman, F., and Stewart, J.W. (1975). The use of iso-1-cytochrome c mutants of yeast for elucidating the nucleotide sequences that govern initiation of translation. In: Organization and Expression of the Eukaryotic Genome: Biochemical Mechanisms of Differentiation in Prokaryotes and Eukaryotes, G. Bernardi, and F. Gros, eds. (New York: Elsevier), pp. 175–191.
Sonenberg, N., Hershey, J.W.B., and Mathews, M.B., eds. (2000). Translational Control of Gene Expression (Cold Spring Harbor: Cold Spring Harbor Laboratory Press).
Sood, R., Porter, A., Ma, K., Quilliam, L., and Wek, R. (2000). Pancreatic eukaryotic initiation factor-2 [x03B1] kinase (PEK) homologues in humans, Drosophila melanogaster and Caenorhabditis elegans that mediate translational control in response to endoplasmic reticulum stress. Biochem. J. 346, 281–293. Abstract Article
Stachelska, A., Wieczorek, Z., Ruszczynska, K., Stolarski, R., Pietrzak, M., Lamphear, B., Rhoads, R., Darzynkiewicz, E., and Jankowska-Anyszka, M. (2002). Interaction of three Caenorhabditis elegans isoforms of translation initiation factor eIF4E with mono- and trimethylated mRNA 5' cap analogues. Acta. Biochim. Pol. 49, 671–682. Abstract
Stover, N.A., and Steele, R.E. (2001). Trans-spliced leader addition to mRNAs in a cnidarian. Proc. Natl. Acad. Sci. USA 98, 5693–5698. Abstract Article
Styhler, S., Nakamura, A., Swan, A., Suter, B., and Lasko, P. (1998). vasa is required for GURKEN accumulation in the oocyte, and is involved in oocyte differentiation and germline cyst development. Development 125, 1569–1578. Abstract
Tarun, S.Z., and Sachs, A.B. (1996). Association of the yeast poly(A) tail binding protein with translation initiation factor eIF-4G. EMBO J. 15, 7168–7177. Abstract
Thompson, S.R., Goodwin, E.B., and Wickens, M. (2000). Rapid deadenylation and poly(A)-dependent translational repression mediated by the Caenorhabditis elegans tra-2 3' untranslated region in Xenopus embryos. Mol. Cell Biol. 20, 2129–2137. Abstract Article
Tranquilla, T., Cortese, R., Melton, D., and Smith, J. (1982). Sequences of four tRNA genes from Caenorhabditis elegans and the expression of C. elegans tRNALeu (anticodon IAG) in Xenopus oocytes. Nucl. Acids Res. 10, 7919–7934. Abstract
Trutschl, M., Dinkova, T.D., and Rhoads, R.E. (2005). Application of machine learning and visualization of heterogeneous datasets to uncover relationships between translation and developmental stage expression of C. elegans mRNAs. Physiol. Genomics 21, 264–273. Abstract Article
Van Doren, K., and Hirsh, D. (1990). mRNAs that mature through trans-splicing in Caenorhabditis elegans have a trimethylguanosine cap at their 5' termini. Mol. Cell Biol. 10, 1769–1772. Abstract
Vellai, T., Takacs-Vellai, K., Zhang, Y., Kovacs, A.L., Orosz, L., and Muller, F. (2003). Genetics: influence of TOR kinase on lifespan in C. elegans. Nature 426, 620. Abstract Article
Wakiyama, M., Imataka, H., and Sonenberg, N. (2000). Interaction of eIF4G with poly(A)-binding protein stimulates translation and is critical for Xenopus oocyte maturation. Curr. Biol. 10, 1147–1150. Abstract Article
Wang, L., Eckmann, C., Kadyk, L., Wickens, M., and Kimble, J. (2002). A regulatory cytoplasmic poly(A) polymerase in Caenorhabditis elegans. Nature 419, 312–316. Abstract Article
Wightman, B.C., Ha, I., and Ruvkun, G.B. (1993). Posttranscriptional regulation of the heterochronic gene lin-14 by lin-4 mediates temporal pattern formation in C. elegans. Cell 75, 855–862. Abstract Article
Zhang, K., and Kaufman, R.J. (2004). Signaling the unfolded protein response from the endoplasmic reticulum. J. Biol. Chem. 279, 25935–25938. Abstract Article
*Edited by Thomas Blumenthal. Last revised January 12, 2005. Published January 28, 2006. This chapter should be cited as: Rhoads, R. E. et al. Mechanism and regulation of translation in C. elegans (January 28, 2006), WormBook, ed. The C. elegans Research Community, WormBook, doi/10.1895/wormbook.1.63.1, http://www.wormbook.org.
Copyright: © 2006 Robert E. Rhoads et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
§To whom correspondence should be addressed. E-mail: [email protected]
†Current address: Departamento de Bioquimica, Facultad de Quimica, Universidad Nacional Autonoma de Mexico, 04510, Mexico DF, Mexico
 All WormBook content, except where otherwise noted, is licensed under a Creative Commons Attribution License.
All WormBook content, except where otherwise noted, is licensed under a Creative Commons Attribution License.