Embryo series courtesy of Einhard Schierenberg
Embryo series courtesy of Einhard SchierenbergTable of Contents
Abstract
The pharynx is a neuromuscular pump at the anterior end of the alimentary tract. It is made up of 20 muscle cells, 20 neurons, and 20 other cells. Pharyngeal activity correlates with food intake. The proper feeding rate, as well as the precise timing of pharyngeal movements, is required for efficient feeding and likely for survival in nature. For most purposes, pharyngeal behavioral analysis requires no more than a routine stereomicroscope and a pair of eyes, but accuracy can be increased by video recording followed by off-line analysis in slow motion. Like other C. elegans behaviors, pharyngeal behavior is sensitive to both the immediate environmental conditions as well as to the history of such conditions.
The movement of bacteria from outside the worm to the intestine is accomplished by two pharyngeal movements: pumps and isthmus peristalses. Pumps consist of coordinated contraction-relaxation cycles of the radially-oriented muscles of the corpus, anterior isthmus, and terminal bulb (Figure 1, 2A and Movie 1). Isthmus peristalsis occurs following one out of every 3-5 pumps and is coupled to the preceding pump (Song and Avery, 2012). During isthmus peristalsis, the lumen of the posterior isthmus opens and closes in an anterior-to-posterior wave (Figure 2B and Movie 1). Peristaltic motion transports bacteria trapped in the anterior isthmus to the grinder in the terminal bulb. The grinder then crushes bacteria so it can be further digested in the intestine (Movie 2). Although isthmus peristalses are coordinated with pumps, it is easiest to study the two motions separately; we dedicate a separate section to each motion. Use of the electropharyngeogram method as a tool to study pharyngeal physiology (Raizen and Avery, 1994) is discussed in the Wormbook chapter Electrophysiological recordings from the pharynx. At the end of this chapter, we include an appendix (Table 2) with a list of useful promoters for restrictive pharyngeal expression of transgenes. We also include in the appendix references for two methods of staining the pharynx (Table 3).
|
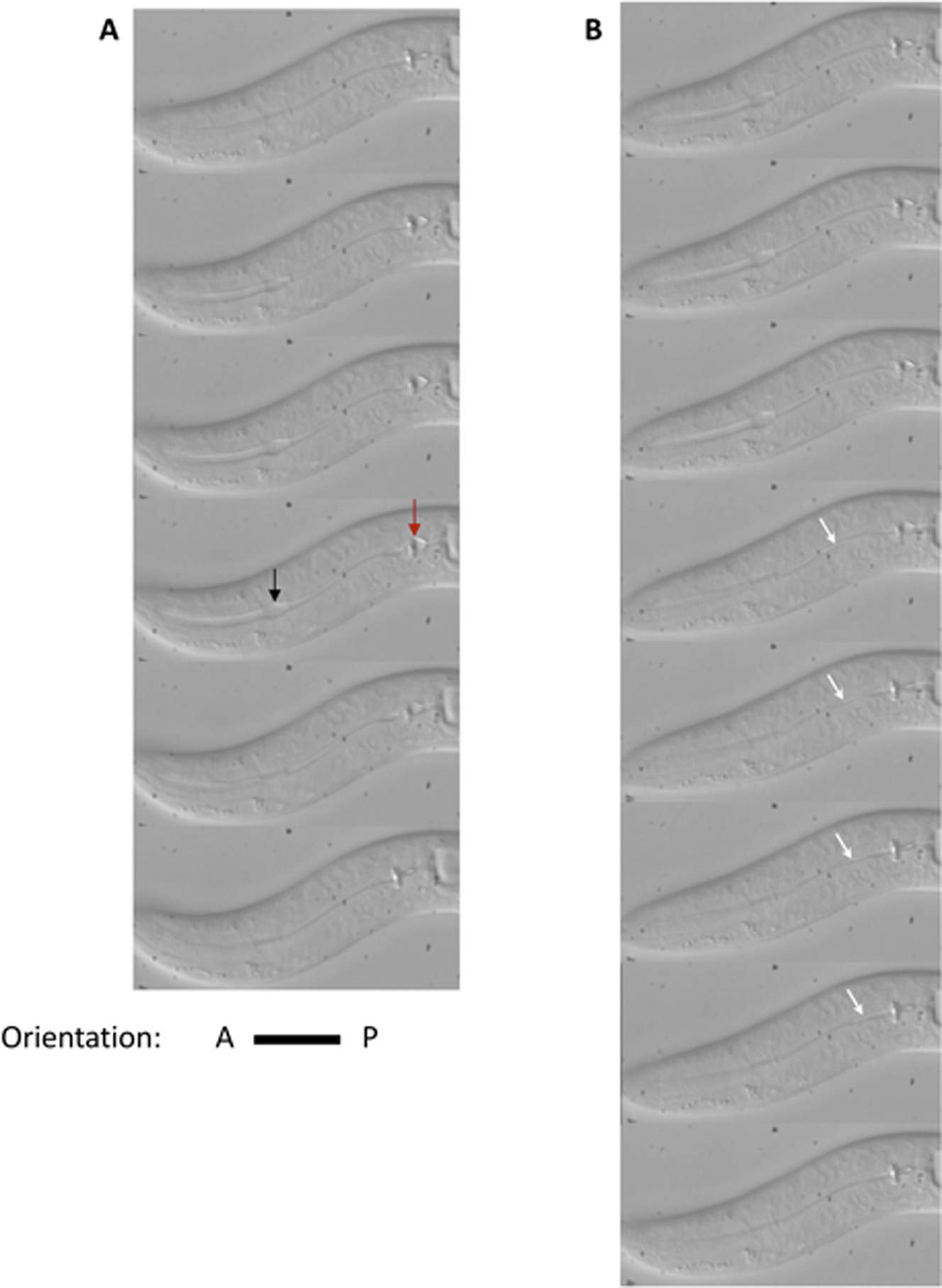
Figure 2: Time lapse images of feeding motion of an L1 animal under a high power DIC compound microscope. (A) Pumping: the corpus (seen as an opening of lumen of the anterior pharynx, black arrow) and the terminal bulb (red arrow) contract and relax synchronously. (B) Isthmus peristalsis: The opening of the lumen of the posterior isthmus is marked with white arrows. During isthmus peristalsis, the posterior isthmus gradually opens and closes in an anterior to the posterior wave.
Several direct and indirect ways of measuring pharyngeal behavior and food intake have been reported (Avery, 1993; Avery and Horvitz, 1990; Fang-Yen et al., 2009; Morck et al., 2004; Song and Avery, 2012). We begin with the simplest and most direct way to study pumping: manual observation.
During feeding, worms suck in and trap bacteria in the corpus, and grind bacteria in the terminal bulb with a structure called the grinder (Figure 1). One complete cycle of synchronous contraction and relaxation of the corpus and the terminal bulb is called a pump (Figure 2A, Movie 1). A simple way to measure feeding is to count how many times worms pump in a minute (pumps per minute: ppm). Because grinder movement in the terminal bulb is easier to see than corpus movement, and because contraction/relaxation cycles are synchronized along the whole pharynx, pumping rate can be measured by counting grinder movements. A stereomicroscope used for daily maintenance of worms (examples: Wild M5A, Leica MS5, and Zeiss Stemi 2000) can be used to count pumps at a magnification of 40-50X (Movie 2). Higher magnifications are required for counting pumps of larvae. Observation of pumping in first larval stage worms (L1s) requires > 100X magnification. Because pumping rate can vary from minute-to-minute or even second-to-second (Hobson et al., 2006), it is sometimes desirable to measure ppm of the same worm several times. This is particularly useful when the worm studied is a precious one (for example, an animal on which a laser-ablation operation had been performed); in such a case, pumping rate can be counted for 10-20 seconds every minute for 10 minutes and the median or mean is then used. Another instance where multiple measurements of the same worm are informative is when a perturbation is predicted to affect the variance rather than the absolute rate of pumping (Hobson et al., 2006). If the number of worms is not limiting and the key parameter of interest is the mean pumping rate, then we recommend measuring the behavior of at least 10 worms. Although it is possible to measure pumping rate of a worm among a population of other worms on the same plate, we find that measurements are more reliable and show reduced variance when worms are individually plated. It is important to use age-matched worms for comparisons. Feeding is easiest to visualize in 6-24 hour old adults.
Under certain conditions, the pumping rate is too fast to count in real time or the motions of individual pumps are abnormal and are therefore not clearly discerned. Under these conditions, it is useful to record movies and replay them at one half to one third of the original speed to count pumping. When using a digital camera, the frame rate should be at least 10 frames per second. Under rare conditions when pumping rates greater than 300 per minute (5 Hz) are expected, the digitization rate should be faster, and at least twice the rate of the pumps.
Direct counting using a stereomicroscope can be used to measure pumping rates of the worms under various experimental conditions. Precise ppm counting of worms cultivated on a full lawn of bacteria that is a low quality food source (e.g. OP50 and its derivative DA837) is tricky because of increased locomotive activity of hungry worms. In such cases, we recommend plating a single worm on a bacterial lawn of approximately 5 mm diameter and recording a video. Table 1 shows ppm as a function of food quality or after surgical manipulations.
Although a simple stereomicroscope is adequate for counting pumps based on grinder movements, a compound microscope equipped with DIC optics is necessary for observing movements in the corpus and isthmus or for measuring the speed of contraction or relaxation. Isthmus peristalsis movements can be directly visualized only with high power optics (see below).
Worms are placed on a 2% agar pad with abundant bacteria and 3-5 µl of M9 or NGM buffer. A cover slip is placed on top of the worms. After a 5-15 minute acclimation period, the worms will begin feeding. They can be viewed with a 40X, 63X, or 100X objective lens equipped with DIC optics. For prolonged viewing of the same worm, it is necessary to move the stage to keep the worm (or worm's pharynx) in the field of view. This is difficult to do in adult worms because their movements are too large to easily track by hand. There are a couple of tricks to minimize movements of large worms while observing feeding under high power magnification. The first is to use worms with a mutation that impairs locomotion. We commonly use worms with mutations in the body muscle levamisole receptor subunit unc-29 (Avery, 1993). Severe paralysis of body muscle, as seen with unc-54 mutants, is best to avoid since it appears to interfere with normal feeding behavior (David Raizen, unpublished observations).
A second recently described trick for monitoring feeding behavior while minimizing locomotion is to surround the worm with a suspension of 0.05-0.1 µm polystyrene beads (Polyscience Corp, Warrington PA) on a high concentration (10%) agar pad beneath a cover slip (Fang-Yen et al., 2012). The increased friction of the beads provides a high barrier to worm body motions and essentially immobilizes the worm. However, the pharynx, which is internal to the worm, continues to behave even when the worm is immobile. A 10-15 minute acclimation period on the pad and in the presence of the beads is required before worms resume pumping, which is stimulated by the addition of serotonin (5 mg/mL) to the pad. When interpreting experiments in which polystyrene beads are used for immobilization, it is important to be aware that mechanical input is known to inhibit pumping (Keane and Avery, 2003).
Under low light conditions, grayscale values of neighboring pixels can be binned to increase camera sensitivity. This will reduce the exposure times of individual frames and allow for faster frame rates. In special cases, for example when measuring the speed of pharyngeal relaxation in subparts of the pharynx, a camera that is specialized for high speed recording (for example, Andor iXon camera) capable of frame rates on the order of 1000 frames per second is required (Fang-Yen et al., 2009).
Using a compound microscope, one can measure the frequency and duration of isthmus peristalsis as well as the relative timing of isthmus peristalses to pumps (Figure 2B, Movie 2). A drawback of this method is that the amount of food given to each worm on the agarose pad cannot be easily controlled and, since food density affects the frequency of isthmus peristalses (Bo-mi Song, unpublished observation), isthmus peristalsis rates on agarose pads may be variable. In contrast to food, it is possible to tightly control the concentration of drugs on the agarose pad. Agarose suspensions are prepared by adding 2% (weight/volume) agarose to an M9 solution containing the drug at the desired final concentration. Avoid using dimethyl sulfoxide (DMSO) as an organic solvent, since at a concentration as low as 0.25% DMSO is an activator of isthmus peristalsis (Bo-mi Song, unpublished observations). Ethanol at concentrations up to 0.25% does not affect either pharyngeal pumping rate or isthmus peristalsis rate, and is therefore a reasonable vehicle for the drugs. Effects of drugs on isthmus peristalsis rate can be examined by placing worms on the drug-containing pads. Isthmus peristalsis is best measured in L1s that are at least three hours old because the pharyngeal muscles of younger worms often twitch. The following is a specific example of the analysis of the effect of a drug (serotonin) on isthmus peristalses. Other drugs can be studied in a similar fashion.
Protocol: Observation of the effect of serotonin on isthmus peristalsis in 3-5 hour old L1 larvae in the absence of food.
After bleach treatment, transfer embryos suspended in 200 µL of M9 onto an unseeded NGM plate and incubate at room temperature for 2 hours. Absorption of the M9 into the agar during the 2-hour incubation will result in unhatched embryos sticking to NGM agar surface, thus facilitating the selective transfer of newly hatched L1s. Transfer L1s within 2 hours of hatching to obtain worms with a defined age.
Transfer the L1s to an unseeded NGM plate and remove unhatched embryos, if any, by aspiration. Incubate the L1s for 3 hours at room temperature until they are 3-5 hour old.
Make a 2% agarose pad that contains 20 mM serotonin. First, make 20 mM serotonin solution by dissolving 0.0232 g of serotonin creatine complex (H7752, Sigma-Aldrich) in 3 mL M9. To completely dissolve the serotonin, vigorous agitation may be required. Then, add 0.06 g agarose to the serotonin solution.
Make an agarose pad on a glass slide using approximately 100 µl of the agarose/serotonin solution. Place 1-3 µl of M9 solution on the pad, transfer 5-10 worms into the solution and put a coverslip on top. After 15 minutes of incubation on the pad, focus attention on the posterior isthmus and count the number of peristaltic movements.
Indirect detection of isthmus peristalsis using fluorescent bacteria or fluorescent latex beads permits the assessment of the effect of food density on the frequency of isthmus peristalsis on an NGM agar surface. During isthmus peristalsis, bacteria and other particles in the corpus pass through the lumen of the posterior isthmus. Bacteria expressing fluorescent proteins can be detected as they pass through the posterior isthmus during a peristalsis (Song and Avery, 2012). When fluorescent latex beads are used for the assay, the ratio between beads and food is important. If the bead concentration is too low, the fluorescent signal will be too low to reliably identify every isthmus peristalsis, whereas if the bead concentration is too high, background fluorescence will obscure the signal from the beads.
Blue light increases isthmus peristalsis rate. Thus, bacteria or beads that emit red fluorescence are preferable since the light used to illuminate red beads is green. We have successfully used 0.5 µm red fluorescent latex beads from Sigma-Aldrich (L3280). The density of food and the age of worms affect isthmus peristalsis rates. We therefore compare isthmus peristalses of equal-age worms. To ensure constant bacterial concentration, we plate equal volumes of bacterial suspension of a defined concentration (defined by optical density at 600 nm) on each plate and incubate for a defined period at the same temperature. To obtain age-matched 1-day-old adults, we transfer L4 animals the previous day.
Protocol: Quantification of isthmus peristalsis rate using mCherry-expressing HB101 or using red fluorescent beads
After a bleach treatment, synchronize the developmental stage of the worms by incubating in 3 mL of M9 for approximately 24 hours at 20-24 °C.
Culture the developmentally synchronized L1s for 54 hours at 20 °C on HB101 until adulthood.
5.5 hours before the assay, seed 10 µl of concentrated mCherry-expressing HB101 culture (OD600=5.0) on NGM plates and incubate at room temperature. When using fluorescent latex beads (L3280, Sigma-Aldrich), mix the beads with HB101 culture (OD600=5.0) at a 1:800 ratio (1 µl bead suspension/800 µl culture).
Remove bacteria from an individual worm by letting it crawl ~1 minute on an unseeded NGM plate. Transfer the worm to the test plate seeded with mCherry-expressing HB101 and observe isthmus peristalsis under a stereomicroscope with a rhodamine fluorescence filter by looking for red signal passing through the posterior isthmus.
Feeding can be indirectly measured using bacteria expressing a fluorescent protein. We commonly use GFP-expressing bacteria. Because worms defecate every 45-50 seconds in the presence of plentiful food (Thomas, 1990), ingested contents stay in the gut only for 5-15 minutes (Leon Avery & Young-Jai You, unpublished data). Therefore, the intensity of fluorescence in the intestinal lumen depends primarily on the intake of food 5-15 minutes prior to observation. It follows that one must be careful when studying feeding by this method under experimental conditions that may affect defecation. An alternative to using GFP bacteria is to use the fluorescent dye BODIPY (You et al., 2008), which has the advantage of assessing feeding over longer time periods (Figure 3). However, BODIPY accumulation can be affected by pumping rate and not only by food ingestion (Young-Jai You, unpublished observations) and therefore should be used with caution when comparing between intake of foods of different qualities or between strains with different efficiencies of food transport.
|
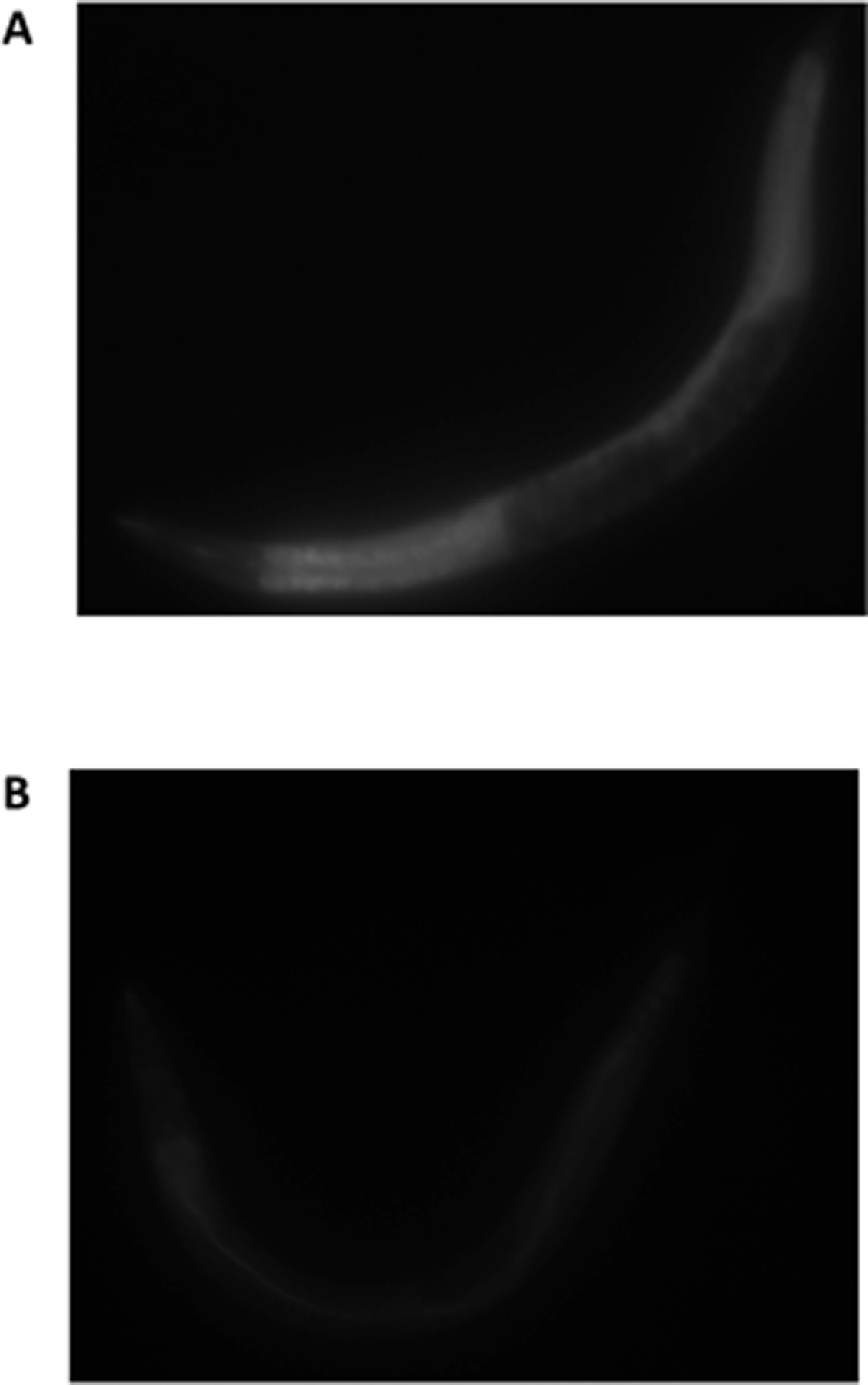
Figure 3: BODIPY staining after 15 minutes of feeding with HB101. (A) Worm was fasted for 12 hours and refed for 15 minutes with HB101 in the presence of BODIPY (B) worm was not fasted but fed with HB101 in the presence of BODIBY for 15 minutes. Reproduced with permission from (You et al., 2008).
Protocol: Using GFP bacteria or BODIPY to measure food intake
Inoculate GFP-expressing bacteria into a 1:1 mixture of LB and M9 to make a bacterial broth and grow overnight at 37 °C without shaking. When kept at 4 °C, the broth can be used for approximately one month.
Maintain both wild type and test strains at 20 °C by picking three adults every day to a new plate seeded with HB101.
Place age-matched individual worms on individual plates seeded with a confluent lawn of GFP-expressing bacteria and allow them to feed ad libitum for the desired period of time. Place a 20 µl drop of 1 M NaN3 on top of each worm to kill it. Treating worms with NaN3 prevents additional feeding while you are transferring the worm to the 2% agar pad and add a cover slip.
Take an image of a whole worm using a GFP filter and a 10X objective lens.
Analyze the fluorescence intensity using digital image analysis software (e.g., Image J)
For BODIPY staining, dissolve the BODIPY dye in DMSO at 1 mg/ml to make a stock solution. Dilute the stock solution and spread on plates to achieve a nominal final concentration of 200 ng/ml (the final concentration of DMSO is 0.02%, which is 10-fold lower than the concentration that can affect isthmus peristalses). After the desired period of feeding, take pictures and analyze the fluorescence intensity using Image J.
Caution: Avoid transferring excessive GFP bacteria, since it will result in high background fluorescence and spoil your analysis.
Note that although NaN3 has been reported to bleach GFP fluorescence expressed in C. elegans cells, it has not been a problem for this method.
Under typical laboratory conditions, worms rarely stop feeding for more than a few seconds unless in lethargus. Lethargus is the ~2 hour period before each of the four molts (Singh and Sulston, 1978). Both pumping and isthmus peristalses cease during lethargus. Cessation of feeding therefore specifically identifies worms in lethargus (Raizen et al., 2008; Van Buskirk and Sternberg, 2007). We use absence of pumping for 10 seconds in the appropriately aged animal to signify lethargus. L3 and L4 lethargus can be readily identified based on absence of pumping under high magnification (40-50X) of a dissecting microscope. To identify L1 and L2 lethargus, higher magnification is needed. We typically use >100X magnification of a stereomicroscope. Unlike the cessation of pumping under conditions of satiety, which is extremely sensitive to environmental conditions (see below), the cessation of pumping during lethargus is a robust behavior that is insensitive to environmental conditions. We have found that even when animals are mechanically stimulated to move continuously for 30 minutes during L4 lethargus, the duration of pumping cessation remains constant at approximately two hours (Robert Driver and David Raizen, unpublished observations). Any pumping observed during lethargus is therefore abnormal and requires non-behavioral anatomical criteria to define the stage as lethargus. For an example of inappropriate pumping during lethargus, see Van Buskirk and Sternberg (Van Buskirk and Sternberg, 2007).
Protocol: Identifying worms within 10 minutes of the start of L4 lethargus:
At the start of the day, transfer 30-50 mid to late L4 worms to a plate fully seeded with bacteria.
Every 10 minutes, observe the plates under 10-20X magnification to search for worms that are not moving. If a worm is not moving, check under 40-50X magnification whether or not it is pumping. If it does not pump for 10 seconds or more, then it has entered L4 lethargus.
Perform experiments on the worms in lethargus. Note that worms that are not pumping always show morphological evidence of the molt under DIC optics including a cap at the anterior end of the buccal cavity.
When worms feed after a period of starvation or when they eat highly-preferred bacteria, they can become quiescent. We call this “satiety quiescence” because it mimics the behavioral sequence of satiety in other animals and because the behavior depends on food quality, nutritional status and the history of feeding. The following detailed protocols are modified from a prior publication (You et al., 2008).
Worms: Maintain both wild type and test strains by picking three adults every day to a new plate seeded with HB101 at 20 °C.
Caution: Contamination with other bacteria can affect feeding and satiety quiescence. We add Streptomycin (200 µg/ml) to plates to prevent growth of other E. coli. Also, since worm growth and satiety quiescence is adversely affected on dry plates (Young-Jai You, unpublished observation), we use plates that are no older than 2 weeks and not dried.
Test plates: Prepare test plates with bacteria from bacterial broth prepared as described above (see protocol: Using GFP bacteria or BODIPY to measure food intake). One day before the assay, seed 35 mm NGM plates with 5 µl of HB101 bacteria broth in the center. Ten plates per test strain are needed. The bacteria lawn needs to be small in order to rapidly find the worms while minimally disturbing them.
Incubate the seeded plates at 37 °C overnight.
After removing the plates from 37 °C, leave them at room temperature for a day before use.
Satiety quiescence can be observed under two conditions: (1) after fasting and subsequent refeeding, and (2) when fed good quality food. The fasting-refeeding condition shows most consistent satiety quiescence.
Day 1:
Synchronization of worms: Transfer 30 L4 hermaphrodites from a non-crowded culture plate (At 20 °C, a plate typically becomes crowded 2-2.5 days after placement of 3 adult wild-type worms on the plate) to a new HB101-seeded plate. L1 synchronization using bleaching doesn't work for obtaining staged worms for satiety quiescence assay for two reasons. First, L1 bleaching and synchronization requires overnight starvation after egg prep. This affects the worms’ metabolism (Young-Jai You, unpublished observation) so it could affect feeding behavior. Second, many mutants grow slower and less synchronized than wild type worms.
Fasting: within 12 hours after transfer, L4 worms become young adults. Transfer young adults individually to 60 mm unseeded NGM plates (i.e., one worm/plate). To minimize the amount of bacteria, transfer worms that are off the bacteria lawn using a minimum amount of bacteria on the worm pick. Tapping the plate enhances worm movement thus increasing the chance that they leave the food. Fast the individually-plated worms at 20 °C for 12 hours.
Day 2:
After 12 hours of fasting, transfer the fasted worms individually to the test plates. Fasted animals should appear pale and bloated with embryos. Transfer 10 worms for each test group. To minimize the difference in refeeding period among test samples, we recommend testing no more than 3 groups, 30 worms in total, at one time.
Place the plates next to the microscope. Do not stack the plates and leave enough space between them to allow you to pick one without touching the others.
Refeed the worms undisturbed for 3 or 6 hours. During this time, minimize vibration and noise around the testing area.
After 3 hours of refeeding, set your microscope magnification to 40 or 50X. Transfer a plate carefully to the microscope base and observe the worm.
The worm should be quiescent, neither moving nor pumping. Start timing to measure the quiescence duration. The quiescence duration is defined as the interval between the beginning of observation of the quiescent animal to the time at which the animal resumes feeding and locomotion.
The first movement a quiescent animal makes is typically a backwards movement, which is followed by resumption of pharyngeal pumping. Occasionally, a worm will make a sub-second body jerk or pump once or twice but then return to being quiescent. Stop the timer only when the worm shows sustained movement and pumping for > 3 seconds.
After measuring the quiescence duration, place the plate back next to the microscope while analyzing the next worm. After an additional 3 hours of refeeding (i.e., 6 hours of refeeding total), measure quiescence duration again.
The phenotype should be scored blind to genotype or other conditions that you are testing.
Caution: quiescence behavior is fragile and sensitive to environmental disturbance. Although we don't know what disturbs worms while we watch them, light, heat and movement of the plates are all candidates.
Maintain worms and prepare test plates as described above. Pick 10 L4s per test group and place each on a test plate seeded with HB101 or Comamonas bacteria. After 18 hours, measure quiescence duration as described above. The phenotype again should be scored blind.
Table 2: Restrictive promoters for driving transgene expression in the pharynx. We list promoters that may be useful for selective expression in the pharynx. Promoters that are expressed in only a single pharyngeal cell type are in bold type.
| Cell | Promoter |
| Pharyngeal muscle pm1-8 | myo-2 (Okkema et al., 1993) |
| Marginal cells mc1-3 | nas-25 (Park et al., 2010) |
| Gland cells g1-2 | nas-12 (Park et al., 2010) |
| M1 neuron | glr-2 (Brockie et al., 2001) |
| M2 neurons | flp-18 (Kim and Li 2004), inx-12 (Altun et al., 2009) |
| M3 neurons | flp-18 (Kim and Li 2004), npr-1 (Coates and de Bono, 2002) |
| M4 neuron | ceh-28 (Pocock and Hobert, 2010; Ray et al., 2008), ser-7b (Hobson et al., 2003) |
| M5 neuron | flp-1 (Kim and Li 2004) |
| I1 neurons | flp-6 (Kim and Li 2004) |
| I2 neurons | flp-15 (Kim and Li, 2004) |
| I3 neuron | inx-9 (Altun et al., 2009) |
| I4 neuron | flp-6 (Kim and Li, 2004) |
| I5 neuron | flp-2 (Kim and Li, 2004), flp-4 (Kim and Li, 2004) |
| I6 neuron | flp-4 (Kim and Li, 2004) |
| MI neuron | sams-5 (Nakano et al., 2010) |
| MC neurons | flp-2 (Kim and Li, 2004), inx-7 (Altun et al., 2009) |
| NSM neurons | tph-1 (Sze et al., 2002), flp-4 (Kim and Li, 2004) |
Table 3: Immunohistochemistry of the pharynx.
| Stained Cells or Tissues | Immunohistochemistry Procedures | Antibody (Dilution rate) | |
|---|---|---|---|
| Serotonin | NSM, ADF, RIH and AIM, (HSN and VC4-5 in hermaphrodite (Sze et al., 2000)/ CP1-6, RPAG , R1Bs, R3Bs, R9Bs, CA1-4 in males (Loer and Kenyon, 1993) | http://home.sandiego.edu/~cloer/loerlab/anti5htlong.html | anti-serotonin rabbit IGG: S5545, Sigma-Aldrich (1:100); Cy-3 conjugated donkey anti-rabbit IGG: 711-165-152, Jackson ImmunoResearch (1:200) |
| Chitin | The buccal capsule, lumen walls of the pharynx and the grinder (Zhang et al., 2005) | http://www.wormatlas.org/images/finneyruvkun.pdf | Fluorescene or Rhodamine-conjugated Chitin-binding Probe, New England Biolabs, Inc. (1:100) |
We thank Leon Avery for comments. D.M.R is supported by R01 NS064030-01A1 and by NARSAD, B-M S. is supported by HL46154 from the US public Health Service and Y-J. Y. is supported by 09SDG2150070 from the American Heart Association.
Altun, Z. F., Chen, B., Wang, Z. W., and Hall, D. H. (2009). High resolution map of Caenorhabditis elegans gap junction proteins. Dev. Dyn. 238, 1936-1950. Abstract Article
Aspock, G., Ruvkun, G., and Burglin, T. R. (2003). The Ceanorhabditis elegans ems class homeobox gene ceh-2 is required for M3 pharynx motoneuron function. Development 130, 3369-3378. Article
Avery, L. (1993). Motor neuron M3 controls pharyngeal muscle relaxation timing in Ceanorhabditis elegans. J. Exp. Biol. 175, 283-297. Abstract
Avery, L., and Horvitz, H. R. (1990). Effects of starvation and neuroactive drugs on feeding in Ceanorhabditis elegans. J. Exp. Zool. 253, 263-270. Abstract Article
Brockie, P. J., Madsen, D. M., Zheng, Y., Mellem, J., and Maricq, A. V. (2001). Differential expression of glutamate receptor subunits in the nervous system of Caenorhabditis elegans and their regulation by the homeodomain protein UNC-42. J. Neurosci. 21, 1510-1522. Abstract
Coates, J. C., and de Bono, M. (2002). Antagonistic pathways in neurons exposed to body fluid regulate social feeding in Caenorhabditis elegans. Nature 419, 925-929. Abstract Article
Fang-Yen, C., Avery, L., and Samuel, A. D. (2009). Two size-selective mechanisms specifically trap bacteria-sized food particles in Ceanorhabditis elegans. Proc. Natl. Acad. Sci. U. S. A. 106, 20093-20096. Abstract
Fang-Yen, C., Gabel, C. V., Samuel, A. D., Bargmann, C. I., and Avery, L. (2012). Laser microsurgery in Ceanorhabditis elegans. Methods Cell Biol. 107, 177-206. Abstract
Hobson, R. J., Geng, J., Gray, A. D., and Komuniecki, R. W. (2003). SER-7b, a constitutively active Gαs coupled 5-HT7-like receptor expressed in the Ceanorhabditis elegans M4 pharyngeal motorneuron. J. Neurochem. 87, 22-29. Abstract Article
Hobson, R. J., Hapiak, V. M., Xiao, H., Buehrer, K. L., Komuniecki, P. R., and Komuniecki, R. W. (2006). SER-7, a Ceanorhabditis elegans 5-HT7-like receptor, is essential for the 5-HT stimulation of pharyngeal pumping and egg laying. Genetics 172, 159-169. Abstract Article
Keane, J., and Avery, L. (2003). Mechanosensory inputs influence Ceanorhabditis elegans pharyngeal activity via ivermectin sensitivity genes. Genetics 164, 153-162. Abstract
Kim, K., and Li, C. (2004). Expression and regulation of an FMRFamide-related neuropeptide gene family in Ceanorhabditis elegans. J. Comp. Neurol. 475, 540-550. Abstract Article
Loer, C. M., and Kenyon, C. J. (1993). Serotonin-deficient mutants and male mating behavior in the nematode Ceanorhabditis elegans. J. Neurosci. 13, 5407-5417. Abstract
Morck, C., Rauthan, M., Wagberg, F., and Pilon, M. (2004). pha-2 encodes the C. elegans ortholog of the homeodomain protein HEX and is required for the formation of the pharyngeal isthmus. Dev. Biol. 272, 403-418. Abstract Article
Nakano, S., Ellis, R. E., and Horvitz, H. R. (2010). Otx-dependent expression of proneural bHLH genes establishes a neuronal bilateral asymmetry in C. elegans. Development 137, 4017-4027. Abstract Article
Okkema, P. G., Harrison, S. W., Plunger, V., Aryana, A., and Fire, A. (1993). Sequence requirements for myosin gene expression and regulation in Ceanorhabditis elegans. Genetics 135, 385-404. Abstract
Park, J. O., Pan, J., Mohrlen, F., Schupp, M. O., Johnsen, R., Baillie, D. L., Zapf, R., Moerman, D. G., and Hutter, H. (2010). Characterization of the astacin family of metalloproteases in C. elegans. BMC Dev. Biol. 10, 14. Abstract Article
Pocock, R., and Hobert, O. (2010). Hypoxia activates a latent circuit for processing gustatory information in C. elegans. Nat. Neurosci. 13, 610-614. Abstract Article
Raizen, D. M., and Avery, L. (1994). Electrical activity and behavior in the pharynx of Ceanorhabditis elegans. Neuron 12, 483-495. Abstract Article
Raizen, D. M., Zimmerman, J. E., Maycock, M. H., Ta, U. D., You, Y. J., Sundaram, M. V., and Pack, A. I. (2008). Lethargus is a Ceanorhabditis elegans sleep-like state. Nature 451, 569-572. Abstract Article
Ray, P., Schnabel, R., and Okkema, P. G. (2008). Behavioral and synaptic defects in C. elegans lacking the NK-2 homeobox gene ceh-28. Developmental neurobiology 68, 421-433. Abstract Article
Singh, R. N., and Sulston, J. E. (1978). Some observations on moulting in Ceanorhabditis elegans. Nematologica 24, 63-71. Article
Song, B. M., and Avery, L. (2012). Serotonin activates overall feeding by activating two separate neural pathways in Ceanorhabditis elegans. J. Neurosci. 32, 1920-1931. Abstract Article
Sze, J. Y., Victor, M., Loer, C., Shi, Y., and Ruvkun, G. (2000). Food and metabolic signalling defects in a Ceanorhabditis elegans serotonin-synthesis mutant. Nature 403, 560-564. Abstract Article
Sze, J. Y., Zhang, S., Li, J., and Ruvkun, G. (2002). The C. elegans POU-domain transcription factor UNC-86 regulates the tph-1 tryptophan hydroxylase gene and neurite outgrowth in specific serotonergic neurons. Development 129, 3901-3911. Abstract
Thomas, J. H. (1990). Genetic analysis of defecation in Ceanorhabditis elegans. Genetics 124, 855-872. Abstract
Van Buskirk, C., and Sternberg, P. W. (2007). Epidermal growth factor signaling induces behavioral quiescence in Ceanorhabditis elegans. Nat. Neurosci. 10, 1300-1307. Abstract Article
*Edited by Oliver Hobert. Last revised April 30, 2012, Published December 18, 2012. This chapter should be cited as: Raizen D., Song B., Trojanowski N. and You Y. Methods for measuring pharyngeal behaviors (December 18, 2012), WormBook, ed. The C. elegans Research Community, WormBook, doi/10.1895/wormbook.1.154.1, http://www.wormbook.org.
Copyright: © 2012 David Raizen et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
§To whom correspondence should be addressed. E-mail: Young-Jai You ([email protected])
 All WormBook content, except where otherwise noted, is licensed under a Creative Commons Attribution License.
All WormBook content, except where otherwise noted, is licensed under a Creative Commons Attribution License.