Embryo series courtesy of Einhard Schierenberg
Embryo series courtesy of Einhard SchierenbergTable of Contents
Abstract
The features that differentiate the C. elegans male from the hermaphrodite arise during postembryonic development. The major male mating structures, consisting of the blunt tail with fan and rays, the hook, the spicules and proctodeum, and the thin body, form just before the last larval molt. Male and hermaphrodite embryogenesis are similar but some essential male cell fates are already established at hatching. The male mating structures arise from three important sets of male-specific blast cells. These cells generate a total of 205 male-specific somatic cells, including 89 neurons, 36 neuronal support cells, 41 muscles, 23 cells involved in differentiating the hindgut, and 16 hypodermal cells associated with mating structures. Genetic and molecular studies have identified many genes required for male development, most of which also function in the hermaphrodite. Cell-cell interactions play a role in patterning all three of the generative tissues. Male-specific neurons, including sensory neurons of the rays, hook, post-cloacal sensilla, and spicules, differentiate at the end of the last larval stage and send out axons to make connections into the existing neuropil, greatly enlarging the posterior ganglia. The hindgut is highly differentiated to accommodate the spicules and the joining of the reproductive tract to the cloaca. A complex male-specific program generates many new muscles for copulation. The cell lineage and genetic program that gives rise to the one-armed male gonad appears to be a variation on that of the hermaphrodite.
An L1 larval male just hatched from the egg is not easily distinguished from a hermaphrodite of the same age. Yet by the time it is a mature adult it will be a very different animal. Not only will it have a radically distinct male gonad full of sperm joined to a posterior cloaca rather than a midbody vulva, it will also have a smaller, thinner body, a tail differentiated for copulation, new muscles, and an augmented and modified nervous system that gives it a whole new set of behaviors (Figure 1). These differences arise because the developmental events that occur postembryonically are extensively different in the male and hermaphrodite. In some tissues, cells that divide postembryonically (blast cells) do so in only one of the two sexes, giving rise to sex-specific cell lineages and differentiated cells. In other tissues, identical blast cells follow different cell lineages and generate different sets of progeny cells. Some already terminally differentiated cells reprogram significantly. Finally, in a matter of hours at the end of the last larval stage, dramatic morphogenetic processes reshape the male tail and generate the hypodermal mating structures.
The detailed cellular architecture of the adult male and its postembryonic cell lineages and development were described by Sulston and coworkers (Sulston et al., 1980). In this work, the lineal ancestry of each non-gonadal cell was identified from reconstruction of lineaged animals. The lineage and fates of cells in the male gonad were determined by Kimble and Hirsh (1979). These papers are the starting points for any survey of C. elegans male anatomy or development.
In the present chapter, the results of studies on the genetic, cellular, and morphogenetic basis for development of each structure or feature, in so far as they are known, are summarized. These studies have revealed that male and hermaphrodite development involve largely the same set of genes. Some genes required for male development have been identified in genetic screens for defective males (many are named mab for male abnormal; e.g., Hodgkin, 1983). Almost all of these genes also have functions in the hermaphrodite. The exceptions are the DM domain transcription factor genes mab-3 and mab-23, which appear to be dedicated specifically to aspects of male development (Lints and Emmons, 2002; Shen and Hodgkin, 1988; see Somatic sex determination). Many additional genes required for male development have been identified among hermaphrodite developmental genes because of their pleiotropic effects on the male.
Molecular studies of transcription patterns have given a similar result: only a small proportion of the genes are enriched in males. Whole-genome microarray analysis found that 4% of the genes were enriched greater than 2-fold in the male soma as compared to the hermaphrodite (Jiang et al., 2001). Of these, 14 were transcription factors that might play a more significant role in male development. Overall it appears that the difference between male and hermaphrodite development arises by the differential use of a common set of genes, directed into male-specific or hermaphrodite-specific developmental programs by the sex determination pathway (see Somatic sex determination).
|
Figure 1. Male-specific structures develop postembryonically. (A) The larval male is almost indistinguishable from the hermaphrodite. The external male mating structures differentiate rapidly at the end of the last larval stage. (B) Ventral view of the adult male tail, Nomarski photomicrograph. (C) Sagittal section through the adult male tail showing the three types of canals; reproductive, digestive, and spicule channel, that join at the proctodeal chamber.
The major somatic reproductive structures of the adult male tail are the acellular fan, the rays, the sclerotic, hardened spicules, the specialized proctodeal chamber with its sclerotic, hardened roof (gubernaculum), and the hook (Figure 1). These structures form in the latter half of the L4 larval stage in a rapid morphogenetic episode covering only a few hours before the last larval molt. This period is characterized by a dramatic anterior retraction of the cells of the tail region (Sulston et al., 1980; Figure 2). Simultaneously, the fan is formed and the rays extend. Retraction is accompanied by fusion of the tail tip hypodermal cells, hyp8-11 (Nguyen et al., 1999). As retraction begins, first hyp8-11 and then more anterior hypodermal cells progressively withdraw from the L4 cuticle, which retains its tapered shape, the intervening space becoming filled with fluid. If the L4 cuticle is broken during this process (by making a hole with a laser), retraction stops, suggesting that one morphogenetic force driving the retraction process might be a build up of fluid pressure between the L4 and adult cuticles (unpublished experiments).
The fan, consisting of a fold in the outer layer of the three-layered adult cuticle, forms as the cells of the tail retract. The adult cuticle forms under the L4 cuticle before retraction begins. Over most of the body the inner and outermost layers are attached by struts, but where the fan forms this is apparently not the case. As a consequence, as the future adult body surface pulls away, the inner layer of the adult cuticle withdraws with it, while laterally the outer layer separates and folds to make the fan. During retraction, muscles in the tail, including the sex-specific muscles (oblique muscles, diagonal muscles) pull and twitch, aiding the remodeling process.
|
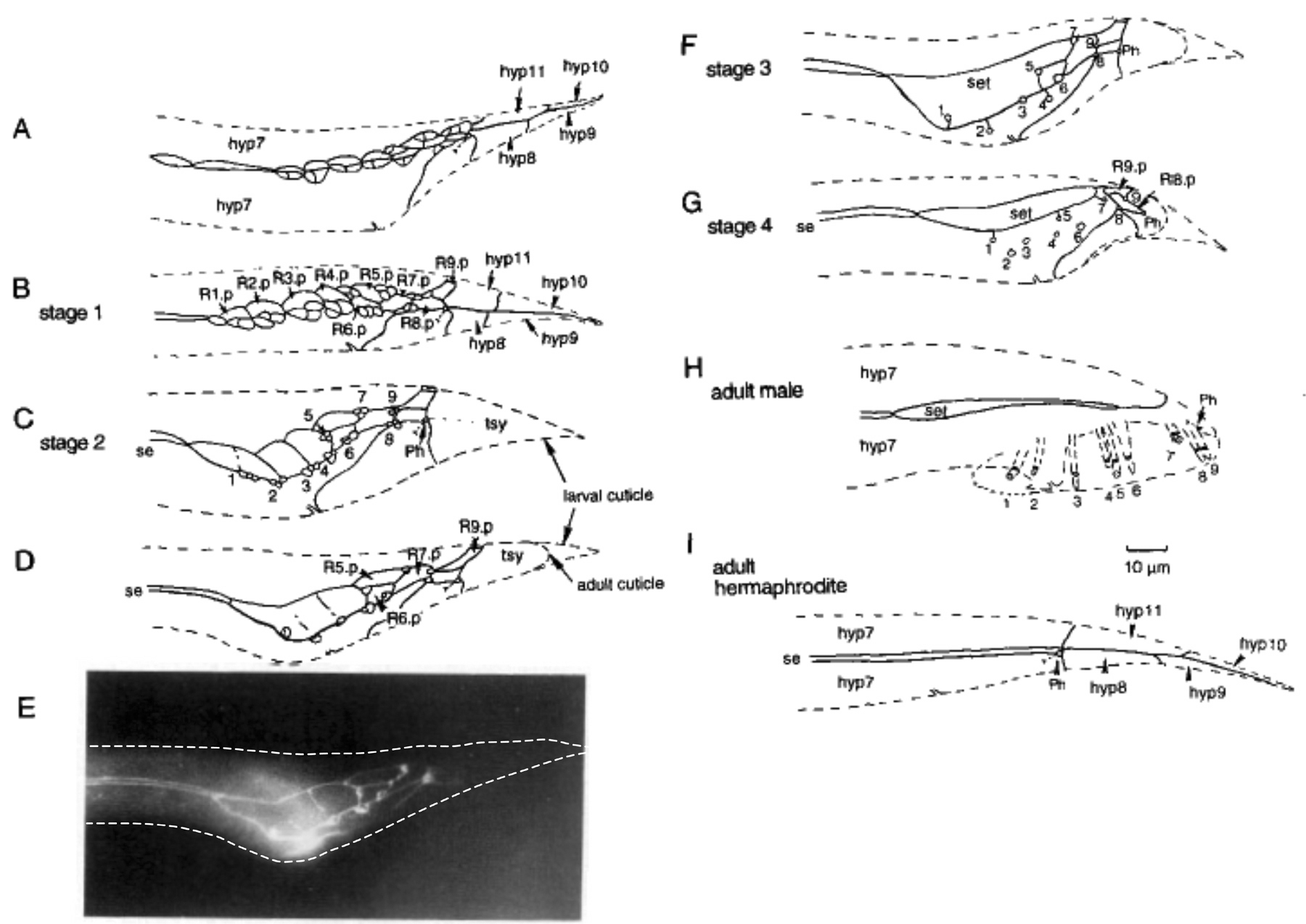
Figure 2. Formation of the fan and rays. The male tail retracts during the last 4 hours of the L4 larval stage (Nomarski photomicrographs). (A) The tail just before retraction begins. (B) The tail tip hypodermal cells, hyp 9 and 10, are the first cells to retract. The arrow indicates the growing fluid-filled cavity between the future adult surface and the L4 cuticle. (C) and (D); the body retracts inside the L4 cuticle, the rays and fan extend. (E) and (F); lateral and ventral views of the adult male tail. In the animal shown in (F); the rays 8 and 9 are fused, as often happens. Reprinted from Developmental Biology, Nguyen et al., 1999, Copyright (1999), with permission from Elsevier.
Hypodermal cells covering the surface of the tail that are likely to be involved in retraction and fan formation are hyp8-11, several cells descended from B.p, and the posterior daughters of the ray precursor cells, the Rn.p cells. The B.p lineage hypodermal cells are arrayed on the ventral side of the L4 tail and constitute a large fraction of the volume of the tail at this time. They retract and withdraw significantly, probably shrinking in volume, during morphogenesis and eventually come to lie above the dorsal roof of the proctodeal chamber, where they form the gubernaculum (see Figure 13; Sulston et al., 1980).
The Rn.p cells, the hypodermal cells generated by the ray sublineage (Figure 7), at first enlarge over the lateral surfaces underlying the region where the fan cuticle will fold out (Figure 3A-C). Possibly they synthesize special cuticle in this region without attachments between the inner and outer layers. Later, R1.p to R5.p fuse to form a separate tail seam (Figure 3D and E), while R6.p to R9.p fuse with hyp 7 (Figure 3F and G; Sulston et al., 1980).
Lep (leptoderan, that is, having a pointed tail) mutants prevent retraction of hyp8-11 while leaving retraction of the other cells unaffected, giving rise to animals with normally-shaped tails except for having long tail tips (Nguyen et al., 1999). A number of so-called leptoderan species have a similar tail phenotype (see The phylogenetic relationships of C. elegans and other Rhabditids). One Lep gene, tlp-1, encodes a C2H2 Zn-finger presumptive transcription factor that may act downstream of a Wnt signal and cell-autonomously promote hyp8-11 retraction (Zhao et al., 2002). Mutations in the posterior Hox gene egl-5 have the opposite effect of Lep mutants: hyp8-11 retract but the remaining tail cells do not and animals have blunt, rounded tails with little or no fan and no rays (Chisholm, 1991). An egl-5 reporter gene has robust expression in tail hypodermal cells for a brief period at the end of L4, suggesting that egl-5 acts cell-autonomously in these cells to promote the retraction process (Teng et al., 2004).
|
Figure 3. The evolving pattern of cells on the lateral surface of the L4 male tail determines where the rays will extend. Cell boundaries in the hypodermal layer have been drawn as visualized by an antibody to a component of adherens junctions. The dashed line is an outline of the body. (A) After the first two divisions of the ray sublineage, each ray precursor cell has generated three cells. hyp7, the large hypodermal syncytium that covers most of the body; hyp8-11, the tail tip hypodermal cells. (B) After the sublineage is completed, the two ray neurons (Rn.aaa and Rn.apa) and structural cell (Rn.app, see Figure 7) cluster against the corresponding Rn.p hypodermal cell. Rn.p is the posterior hypodermal daughter of the ray precursor cell Rn (see Figure 7). (C) The neurons and structural cells have withdrawn leaving attachments at the surface, while the Rn.p cells have enlarged. R1.p and R2.p are beginning to fuse. hyp8-11 have fused to form the tail syncytium (tsy). Ph: phasmid. se: body seam. (D) R1.p to R5.p are almost finished fusing to form the tail seam (set). The neurons have completely withdrawn, leaving only the structural cell attached to the hypodermis. (E) Imunofluorescent photomicrograph of animal shown in D. (F) The structural cells (Rnst) are displaced from the lateral line where the margin of the fan will form. R1st, R5st, and R7st are dorsal of the lateral line, while R2st, R4st and R8st are ventral of it. Rays 3 and 9 remain at the margin. Ray 6 does not open to the exterior. (G) The set retracts after the structural cells become irreversibly attached on the surface. H. Adult male tail. Ph: phasmid. I. Adult hermaphrodite tail for comparison. Reprinted from Fitch and Emmons (1995).
Male and hermaphrodite embryos generate the same set of cells and presumably have the same cell lineages, although the male embryonic cell lineage has not been explicitly determined. Some of the few overt differences between the male and the hermaphrodite hatchee arise by differential programmed cell deaths: in the male, the HSN neuron, which innervates the vulval muscles, dies, whereas in the hermaphrodite, the four CEM head sensory neurons die (Sulston and Horvitz, 1977). In addition, in the hermaphrodite, both left-right pairs of coelomocytes are anterior of the gonad whereas in the male one of two left-side coelomocytes is posterior of the gonad primordium (Sulston and Horvitz, 1977). Finally, the fates of some of the major blast cells appear to have been established by the end of embryogenesis. This is most obvious for the rectal epithelial cell B, which is enlarged in the male. Furthermore, although cellular anatomy of male and hermaphrodite L1 larvae is very similar, comparisons at the ultrastructural level have not been made. In particular, the connectivity of the nervous system in the adult male and in the L1 of either sex has not been determined and so it is not known to what extent the wiring established during embryogenesis may differ between the sexes.
The mating structures of the male tail arise postembryonically from three sets of dividing cells (blast cells). The rays are from blast cells in the lateral hypodermis (V5, V6, T), the hook is from blast cells in the ventral hypodermis (P10, P11), and the spicules and highly differentiated cloacal canal are from blast cells in the rectal epithelium (B, F, U, Y; Figure 4). The identities of these blast cells are in part fixed during embryogenesis and in part determined through signaling events that occur postembryonically. However, no major inducer or inducing tissue (such as the gonad), of male-specific tail development or of any male structure has been identified analogous to the anchor cell in development of the hermaphrodite vulva (see Vulval development; Sulston et al., 1980; Sulston and White, 1980). In the male tail, signals appear to come from multiple and sometimes redundant sources. The Hox genes mab-5 (ortholog of Drosophila ftz) and egl-5 (ortholog of Drosophila AbdominalB) play important roles in patterning all three of these generative tissues.
|
Figure 4. Origins of the major hypodermal mating structures during postembryonic development. The locations of the blast cells in the posterior of an L1 larval male are shown (top: sagittal view; bottom: lateral view). Ventral neuroectoblasts P10 and P11 give rise to the hook and hook sensillum; rectal epithelial cells B and Y give rise respectively to the internal spicules and the post-cloacal sensilla; lateral seam cells V5, V6 and T each give rise to a subset of the rays.
The nine pairs of rays are generated by the three most posterior pairs of seam cells, V5, V6, and T (Figure 4). Though the rays are similar to each other in structure, each also has unique properties (see below). Hence each of these blast cells expresses a different developmental program.
T is preprogrammed to give rise to rays 7, 8, and 9 (Figure 4). Asymmetric cell divisions in the T lineage require Wnt signaling. Correct orientation of cell divisions is dependent on expression of a Wnt ligand (LIN-44) from the posteriormost hypodermal tail tip cells, hyp8-11 (Herman et al., 1995; see Wnt signaling). Hypomorphic alleles of the gene mab-19 specifically disrupt the T lineage and result in loss of the T rays (Sutherlin and Emmons, 1994).
Anterior of T all the way to the head, all the seam cells are to some degree equivalent to one another. All of them have a propensity to generate rays, but for V1-V4 this propensity is inhibited through a not well understood mechanism activated by contacts with their seam cell neighbor (Austin and Kenyon, 1994; Sulston and White, 1980). Inhibition requires the hairy homolog LIN-22 and involves a Wnt pathway, which both promotes ray formation and orients cell division in the seam (Maloof et al., 1999; Wrischnik and Kenyon, 1997; see Wnt signaling). The inhibitory pathway prevents expression of the Hox gene mab-5, which promotes ray development (Hunter et al., 1999). There is a directional bias to the signaling system: after a cell is experimentally removed it is the anterior neighbor that replaces it (Sulston and White, 1980).
V5 and V6 are preprogrammed to overcome this cell contact-mediated inhibition of ray formation. V6 overcomes the inhibitory effects of its neighbors by expressing the homeobox transcription factor PAL-1, an ortholog of Drosophila caudal (Hunter et al., 1999; Waring and Kenyon, 1990). Genes required for establishing V5 identity remain unidentified. mab-5 and egl-5 are essential for the V6 lineage program, and mab-5 is essential for V5 (Chisholm, 1991; Kenyon, 1986). In V6, mab-5 is activated by pal-1 during mid-embryogenesis (Hunter et al., 1999). mab-5 is necessary for activation of egl-5 in certain lineage branches, giving rise to some of the differential properties of the rays (see below, Figure 9) and also for activation of the proneural bHLH transcription factor gene lin-32, which activates the ray sublineage itself (Ferreira et al., 1999; Kenyon, 1986; Zhao and Emmons, 1995). In the V5 lineage, the expression of mab-5 cycles on and off repeatedly to regulate cell proliferation, ray formation, and ray identity (Salser and Kenyon, 1996).
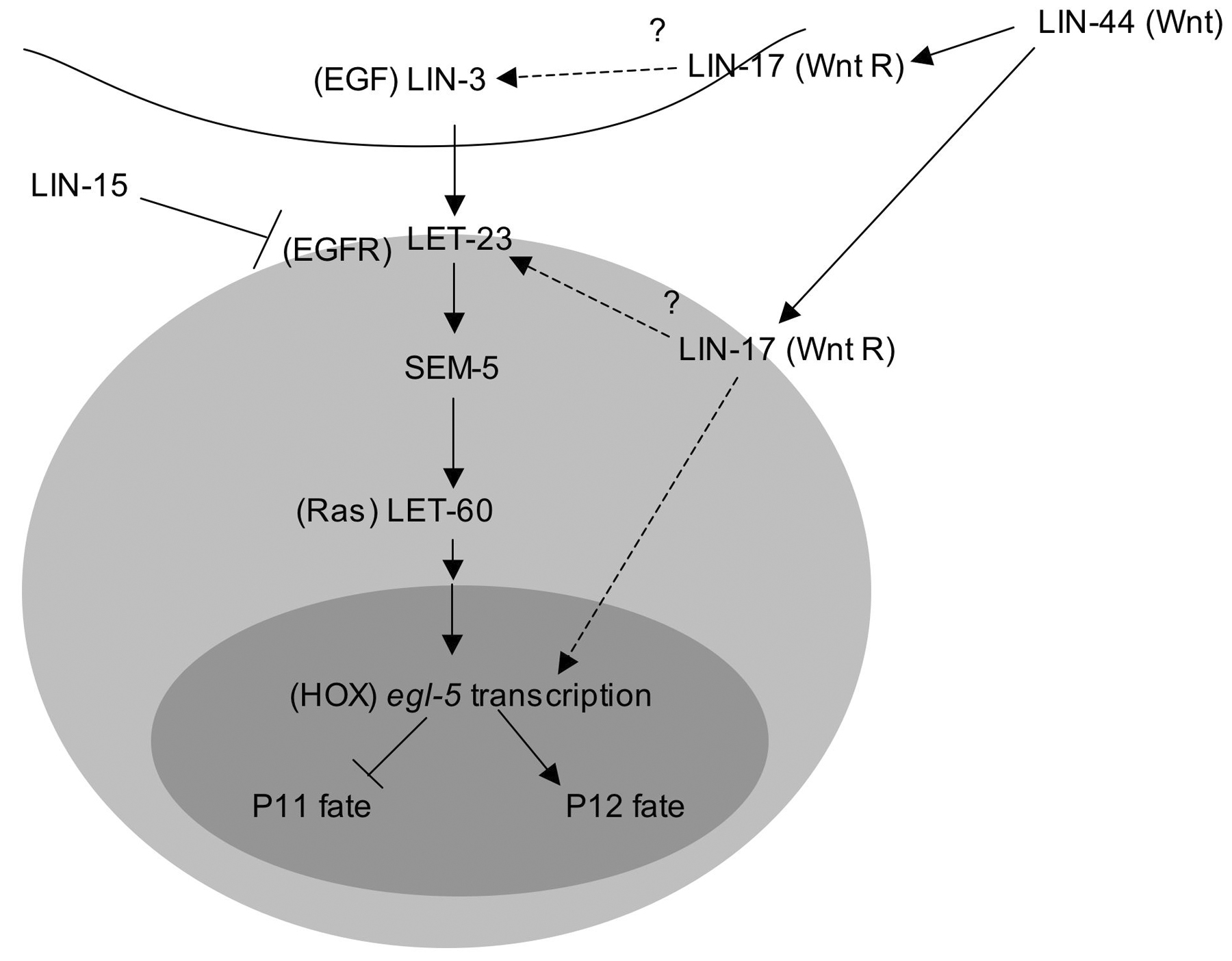
Patterning within the ventral hypodermis that gives rise to the hook is established by the Hox genes. mab-5 is initially expressed in P7-P12 and remains on throughout postembryonic development in the descendants of P7-P11 (Salser et al., 1993). Expression begins during mid-embryogenesis, shortly after these cells are born (Cowing and Kenyon, 1992). Expression of egl-5 is activated in P12 descendants during L1 by an EGF signal (LIN-3/LET-23/LET-60 pathway) and a Wnt pathway signal (LIN-44/LIN-17) and turns off mab-5 expression (Figure 5; Ferreira et al., 1999; Jiang and Sternberg, 1998). Thus, mab-5 and egl-5 are mutually repressing: in egl-5 mutants, mab-5 remains on in P12 descendants and in mab-5 mutants, egl-5 comes on in P10 and P11 descendants. These expression patterns correlate well with the genetic requirements for mab-5 and egl-5 in specifying the fates of the respective cell sublineages (Chisholm, 1991; Kenyon, 1986; Salser et al., 1993).
Epidermal blast cell B, which is a greatly enlarged and easily identifiable cell lying posterior of the rectum in the L1 male, is the major contributor to the internal structures of the male tail, giving rise to 42 cells that differentiate into the spicules, proctodeal chamber, and many neurons, including two of the sensory neurons of the post-cloacal sensilla. The remaining cells of the post-cloacal sensilla as well as an interneuron come from Y (11 progeny). F gives rise to two types of interneurons plus rectal epithelium (8 progeny) and U gives neurons similar in type to those from F and two rectal epithelial cells involved in joining the vas deferens to the cloaca (6 progeny). The locations of these cells around the rectum are shown in Figure 4.
|
Figure 5. EGF and Wnt signals cooperate to induce expression of Hox gene egl-5 in P12.p. egl-5 inhibits mab-5 expression and is sufficient to establish P12.p fate. The sources of the ligands have not been established. Reprinted with permission from Jiang and Sternberg, 1998, Company of Biologists Ltd.
The fates of these cells, as well as the additional rectal cells K and K', are apparently fixed during embryogenesis (Sulston et al., 1980). F and U are sister cells and B and U are lineal homologs, while Y derives from an unrelated branch of the embryonic cell lineage. All of these cells require egl-5 for correct differentiation, apparently in both sexes (Chisholm, 1991). In egl-5 mutant males, the cell lineages are very abnormal and none of the structures normally derived from them, including the spicules, proctodeum, and post-cloacal sensilla, are made.
In addition to egl-5, three transcription factors have been identified that contribute to specifying the fates of the rectal epithelial cells. In mutants of the Tbox transcription factor gene mab-9, B and F are transformed to Y and U, respectively (Chisholm and Hodgkin, 1989; Woollard and Hodgkin, 2000). In mutants of the PAX type transcription factor gene egl-38, F is transformed to B and U is transformed to Y (Chamberlin et al., 1997). In mutants for the C2H2-type Zn finger transcription factor gene lin-48, U is transformed to B (Chamberlin et al., 1999). lin-48 is a direct transcriptional target of EGL-38 (Johnson et al., 2001). Finally, in mutants of the cell signaling gene lin-12, Y takes the fate of its sister cell, the DA9 motor neuron (Greenwald et al., 1983).
A Wnt signaling pathway, activated by the LIN-44/Wnt signal from the tail tip hypodermal cells through the LIN-17 receptor, is necessary for the asymmetry of the first division of B, which separates the precursor of the spicules (B.a) from the precursor of ventral hypodermis and dorsal proctodeal hypodermis (which generates the gubernaculum) and a few neurons (B.p; Herman and Horvitz, 1994; Sternberg and Horvitz, 1988). The B.a fate requires the Pax6 transcription factor VAB-3 (Chamberlin and Sternberg, 1995). Further cell-cell interactions and inductions that pattern cell fates later in these lineages are described below.
Among the several male-specific sensilla in the tail, the rays are the most evident (Figure 1). The rays extend during the period of retraction when the fan forms (Figure 2). Each ray consists primarily of a tube of hypodermis (hyp7) inside of which run processes of two sensory neurons and the ray structural cell (Chow et al., 1995; Sulston et al., 1980; Figure 6). At the tip, the structural cell, attached to the hypodermis by adherens junctions, surrounds the two neurons and creates a hole to the exterior. Either the structural cell grows around the neurons and fuses with itself, or the neurons create a hole through the structural cell. Extension of the ray requires the structural cell but what influence the structural cell exerts on the hypodermis and the mechanics of how the hypodermis itself extrudes this tubular element are not known (Chow et al., 1995; Sulston et al., 1980).
Close apposition of the cuticle around each ray requires a class of at least 8 genes in which mutations result in rays with an expanded, lumpy shape (Ram phenotype for ray morphology abnormal; Baird and Emmons, 1990). Three Ram genes, ram-1, ram-2 and ram-4, encode collagen proteins required specifically in the male tail (Y. L. Yu, W. S. Hui, Y. M. Lam and K. L. Chow, unpublished data). Two encode modifying enzymes: a membrane bound thioredoxin-like protein (DPY-11) and a prolyl hydroxylase (DPY-18), both expressed in the hypodermis; these two enzymes are likely responsible for posttranslational modifications of collagens and other matrix molecules synthesized in the hypodermis (Hill et al., 2000; Ko and Chow, 2002; Winter and Page, 2000). In addition, a novel transmembrane protein expressed in the ray structural cell (RAM-5; Yu et al., 2000) and another type II transmembrane signaling molecule acting non-autonomously in the male tail region (MAB-7; S. W. Tsang, C. Q. Nguyen, D. H. Hall and K. L. Chow, unpublished) are both required for cell cell communication events between the structural cells and hypodermis in the male tail. These gene products acting on the cell surface and the matrix likely participate in cross-linking ray cell components and transducing signals among the cells in each sensory ray.
|
Figure 6. Schematic diagrams of the four types of male-specific sensilla in the tail. The cell bodies are located in various ganglia in the tail (not near the sensilla as shown). Arrows indicate the openings to the exterior. Colored cells: neurons; black: support cells (socket and sheath cells); grey: hypodermal cells.
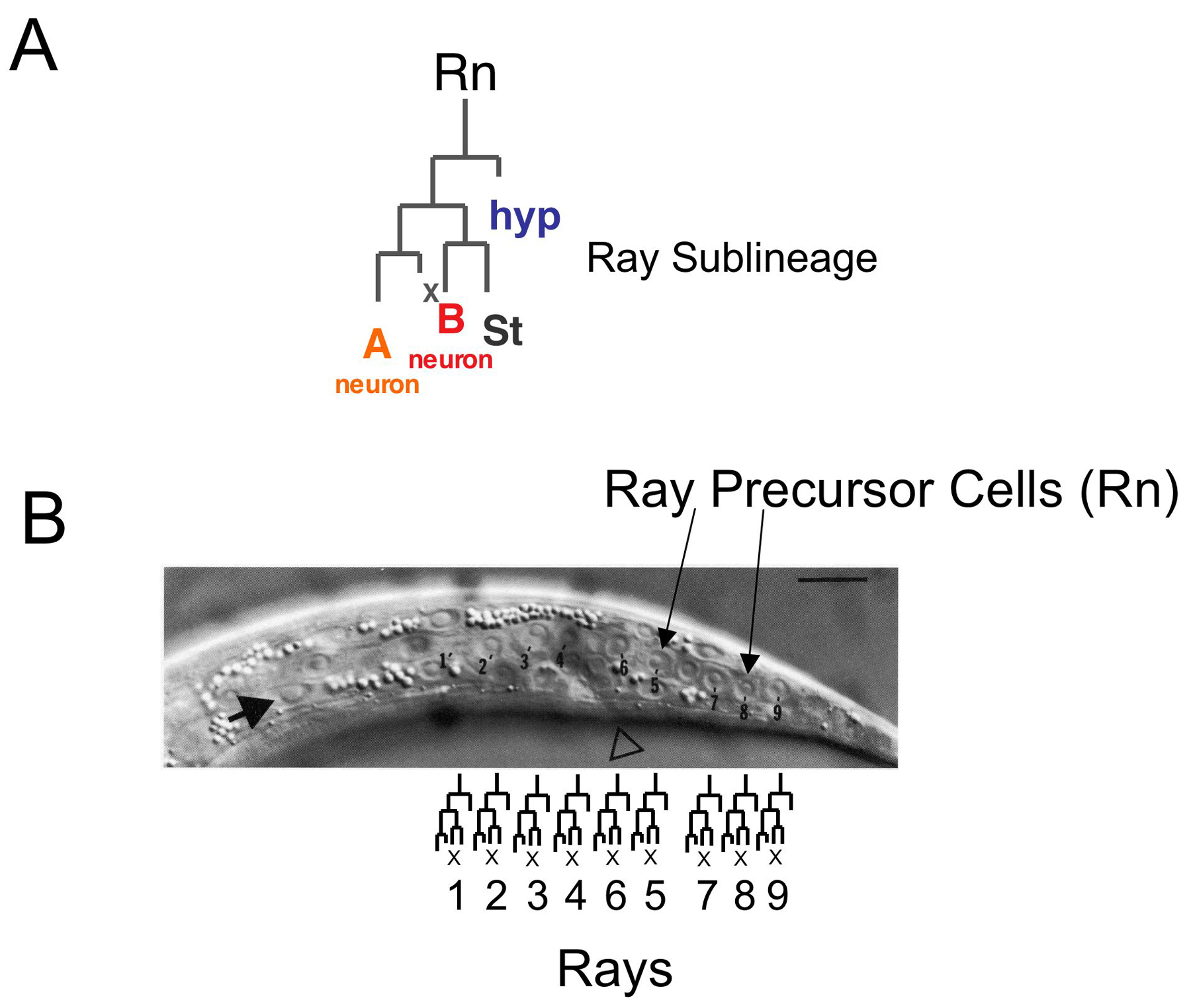
The position where each ray forms is determined by the site where its structural cell attaches to the hypodermis and cuticle. Attachment occurs during the L4 larval stage before retraction begins. The ray cells are born within the hypodermal layer, joined to each other and to hypodermal cells by adherens junctions (Figure 3A and B). Their anteroposterior order in the seam is the same as their final order within the fan except that ray 6 cells are born between ray 4 and ray 5 cells (Figure 7B). Thus, with the exception of ray 6, anteroposterior ray order is determined by the cell lineage.
The spacing and locations of the rays may be determined in part by interactions with the Rn.p cells, the hypodermal cells generated at the first division of the ray sublineage (Figure 7A). Ray cell clusters tend to locate at the junctions of two Rn.p cells, which spread out over the surface in a distinctive pattern (Figure 3C). Ray 5 and 7 cell clusters are born above their respective Rn.p cells; the others are born below. As the neurons and structural cell differentiate, the attachment point of the ray 1 structural cell moves out of the seam into the dorsal hypodermal territory, those of rays 2 and 4 move into the ventral territory, and those of rays 3 and 9 remain in the seam (Figure 3F). As a result of these initial localizations and subsequent displacements, when retraction occurs the tips of rays 1, 5, and 7 open on the dorsal surface of the fan, 2, 4, and 8 open on the ventral surface, and 3 and 9 open at the fan margin (Baird et al., 1991). Ray 6 does not open to the exterior and is localized by being below R6.p, which itself moves ventrally out of the seam.
|
Figure 7. The ray sublineage. (A) The three cells of each ray (RnA: A-type sensory neuron; RnB: B-type sensory neuron; Rnst: ray structural cell) are descended from a single ray precursor cell (Rn). hyp: the Rn.p hypodermal cell. ×: programmed cell death. (B) The row of Rn cells in the L3 seam before they express the ray sublineage (Nomarski photomicrograph). Already by this stage the fates of the Rn cells are established so that each generates specifically one of the nine rays. The open triangle points to the positioning of the cloaca; the arrow indicates a landmark, the hypodermal cell V6.a.
In addition to involving interactions between ray cells and surrounding hypodermal cells, ray formation requires a sorting process involving the ray cells themselves. In this sorting process, each structural cell interacts with and surrounds the neurons belonging to its own ray and rejects interactions with the structural cells and neurons of the other rays. Failure of this system in mutants results in adults with fused rays (Baird et al., 1991). This phenotype indicates that in wild type, the rays express functions that give the cells distinct morphogenetic identities allowing for their proper assembly into individual organs.
The conserved semaphorin, ephrin, and TGFβ signaling pathways are involved in regulating both the interactions between ray cells and hypodermal cells and the interactions among the ray cells themselves. The sensitive response of the ray system to the functioning of the semaphorin and ephrin signaling pathways has allowed an elegant series of studies defining the components of these pathways. Interaction between ray 1 cells and the hypodermal cell R1.p is regulated by the semaphorins semaphorin 1a (SMP-1) and semaphorin 1b (SMP-2) and their corresponding receptor PlexinA (PLX-1; Fujii et al., 2002; Ginzburg et al., 2002). The anteroposterior position of PLX-1-expressing ray 1 is influenced by a semaphorin 1 signal emanating from the hook (Dalpe et al., 2004). Interactions between ray cells are regulated by semaphorin 2a (MAB-20), ephrin-4 (EFN-4/MAB-26), and a TGFβ signal (UNC-129) (Baird et al., 1991; Chin-Sang et al., 2002; Chow and Emmons, 1994; Ikegami et al., 2004; Roy et al., 2000). The ligands MAB-20 and EFN-4 are expressed in most or all the rays (Hahn and Emmons, 2003; Roy et al., 2000). The ray-specificity of their effects may be brought about by the ray-specific expression patterns of UNC-129 and the MAB-20 receptor plexin 2 (PLX-2; Ikegami et al., 2004).
The three cells of each ray descend from a single neuroblast (ray precursor cell) that expresses a stereotyped cell sublineage during the L3 and L4 larval stages (Figure 7; Sulston et al., 1980; Sulston and Horvitz, 1977). Expression of this sublineage requires the proneural bHLH transcription factors LIN-32, an ortholog of Drosophila atonal, and HLH-2, an E/daughterless ortholog, presumably acting as a heterodimer (Portman and Emmons, 2000; Zhao and Emmons, 1995). The DM domain transcription factor mab-3, a member of a conserved class of male-specific regulatory transcription factors, enhances the activities of these two proteins and is required for efficient ray production (Shen and Hodgkin, 1988; Yi et al., 2000). LIN-32 and HLH-2 are required not only for expression of the ray sublineage and neurogenic potential, but also for expression of specific differentiated properties of the ray neurons and support cell (Portman and Emmons, 2000).
Differentiation of the ray cells occurs very rapidly after they are born. They delaminate out of the hypodermal layer leaving behind cell processes as their cell bodies move into the lumbar ganglia (Sulston et al., 1980; Figure 3). From there, ray neuron axons grow through circumferential commissures and enter the preanal ganglion, where they branch and make multiple synaptic contacts (see section 7).
Additional genes expressed in the ray sublineage and in mature ray cells have been identified by microarray experiments, taking advantage of the rayless phenotype of a lin-32 hlh-2 double mutant (Portman and Emmons, 2004). Prominent among differentiation genes expressed in all the rays are a β-tubulin isoform, tbb-4, (A and B-type neurons) and components of the presumptively mechanosensory polycystin signaling pathway, lov-1 and pkd-2 (B-type neurons only), as well as four novel genes (cwp genes) co-expressed with lov-1 and pkd-2 and postulated to constitute additional components of the pathway (Barr et al., 2001; Barr and Sternberg, 1999; Portman and Emmons, 2004). There are many additional genes expressed in subsets of the rays as explained in the next section.
In addition to having distinct morphogenetic identities, as discussed above (see section 5.1.3), the rays also have distinct molecular identities. Although the A-type and B-type neurons are considered to represent two distinct neuron types because of shared ultrastructural features (Sulston et al., 1980), within each type neurons in different rays also have unique features that define molecular subtypes. This property supports the observation that different rays play somewhat differing roles in guiding male copulation (Liu and Sternberg, 1995). Subtype-specific genes include those associated with neurotransmitter synthesis (Figure 8; Lints and Emmons, 1999; Lints et al., 2004; Loer and Kenyon, 1993; Schinkmann and Li, 1992; Sulston et al., 1980) and genes encoding receptors (Portman and Emmons, 2004; Troemel et al., 1995). Subtypes also appear to have subtype-specific patterns of synaptic targets within the lumbar and pre-anal ganglia and hence presumably also differentially express yet to be defined genes associated with axon targeting (SWE, D. Albertson, D. Hall and M. Xu, unpublished observations).
|
Figure 8. Neurotransmitters expressed by the rays. Expression of the monoamine neurotransmitters serotonin and dopamine and of 5 neuropeptide genes tested differs among the rays (Lints et al., 2004).
|
Figure 9. Lineage-specific expression of Hox genes establishes the identities of the V5 and V6 rays. (A) The postembryonic cell lineages that give rise to the rays. Cell fates in these lineages are fixed once the blast cells V5, V6, and T begin to divide. (B) MAB-5 is expressed in an ON/OFF pattern in the V5 lineage (Salser and Kenyon, 1996). Expression must be OFF in V5.p to allow formation of the postdeirid. It must be ON in V5.ppp to induce the equational division of this cell and then OFF again to prevent expression of a ray by the V5.pppa branch. Finally it must be ON in V5.ppppp to generate a ray but go OFF in the sublineage to allow expression of the ray 1 identity. In V6, MAB-5 expression begins during embryogenesis and remains ON throughout postembryonic development except in the ray 3, 5, and 6 branches. (C) EGL-5 expression begins in V6.ppp and in the ray 3 branch (Ferreira et al., 1999).
Ray neuron subtype and ray morphogenetic identity are established coordinately—thus each ray has a unique ray identity (Lints et al., 2004). Ray identities are established by the time the ray precursor cells are born. Determined cell fates are established at each division of the cell lineage, no regulation of cell fates being observed in cell ablation experiments once the V5, V6 and T blast cells begin to divide (Chow and Emmons, 1994; Sulston et al., 1980; Sulston and White, 1980). Critical expression patterns of the Hox genes mab-5 and egl-5 in the ray precursor cells and ray sublineages are set up during this postembryonic lineage period and appear to constitute one central element of the pattern formation mechanism for the V5 and V6 lineages (Figure 9; Ferreira et al., 1999; Salser and Kenyon, 1996). A TGFβ signal further defines the identity of ray 5 (Lints and Emmons, 1999). Patterning of the T rays (rays 7-9) is also affected by a TGFβ signal (Liang et al., 2003; Morita et al., 1999; Savage-Dunn et al., 1996; Suzuki et al., 1999).
Details of a transcription factor code underlying ray identity have been most clearly established for the V6 rays (rays 2-6; Figure 10; Chow and Emmons, 1994; Chow et al., 1995; Lints and Emmons, 1999; Lints et al., 2004; Zhang and Emmons, 1995). For rays 2-6, expression of egl-5 separates non-ray 2 states from a ray 2 ground state. egl-5 alone promotes ray 3 identity. Addition of a signal of the TGFβ family (DBL-1) differentially received by ray 5 elevates this ray to ray 5 identity. Expression of a PAX6 isoform (MAB-18) in ray 6 differentiates this identity from that of other rays. Expression of mab-5 separates ray 2 identity from ray 1 identity (Salser and Kenyon, 1996). Additional regulatory pathways yet to be defined (most likely including the Wnt pathway) are required for a complete model.
The hook, which appears during the process of retraction in L4 lethargus, is a sclerotized point of cuticle formed by the ventral hypodermal cell P10.papp. As the contractions of the diagonal muscles pull the adult cuticle away from the L4 cuticle on the ventral side, the hook is left protruding from the surface. Considerable rearrangements of underlying hypodermal cells that contribute to the structure occur during this process (Sulston et al., 1980).
The nine cells that together form the hook and hook sensillum are born during the L3 larval stage from cell lineages generated by the posterior daughters of the ventral neuroectoblast cells P10 and P11: P11.p contributes three hook-associated hypodermal cells and P10.p generates the cells of the sensillum (the two neurons HOA and HOB, a socket cell and a sheath cell) and the hypodermal cell that generates the hook proper (P10.papp; Sulston et al., 1980; Figure 6). As is the case with the Pn.p cells that generate the hermaphrodite vulva (see Vulval development), the fates of P10.p and P11.p are established by cell signaling within an equivalence group that also includes P9.p. P9.p, P10.p and P11.p fates all require mab-5 (Kenyon, 1986; Salser et al., 1993). These three cells have equal potential and form an equivalence group in which P11.p fate is 1°, P10.p fate is 2°, and P9.p fate (fusion with hyp 7 or division followed by fusion of its daughters) is 3° (Sulston and White, 1980). In equivalence groups, cells of equal potential communicate to allocate separate fates. They do so in such a way that after removal of one cell (in a cell ablation experiment) the remaining cells alter their fates in a hierarchical fashion: 2° replaces 1° and 3° replaces 2°. Patterning within the P9.p-P11.p equivalence group requires genes of the LIN-3/LET-23 and LIN-12 signaling pathways, with LIN-12 defining the 2° fate, as it does in the vulval equivalence group (Greenwald et al., 1983). However, so far, no source of LIN-3/EGF has been identified and details of the patterning mechanism remain unclear (Emmons, 1997; Emmons and Sternberg, 1997).
Specification of the differentiated properties of the neurons has been explored for HOB. A key element of the signal transduction mechanism of HOB, which may involve mechanosensation of the vulval lips, is constituted by polycystin proteins LOV-1 and PKD-2. These proteins are also present in two other classes of male-specific sensory neurons, the RnB neurons of the rays and the CEM neurons (Barr and Sternberg, 1999). The proteins EGL-46, a C2H2-type zinc-finger transcription factor, and EGL-44, a transcription enhancer factor of the TEA domain class, are required for expression of lov-1 and pkd-2 in HOB (but not in RnB; Yu et al., 2003).
Sulston and coworkers (1980) established the structure of the paired sensilla, each containing three sensory neurons (PCA, PCB, and PCC) located just posterior of the cloaca, and the lineal origin of each cell (Figure 6). All are from the Y lineage except PCCL and PCCR, which are from the B lineage. Apart from the studies of genes that are involved in specifying the Y and B cell fates, discussed above (see section 4.4), studies of specification and differentiation of the cells of the post-cloacal sensilla themselves have not been reported.
Each hardened, elongated spicule is composed of two sensory neurons which are exposed at the tip (SPV and SPD), two sheath cells, and four socket cells (Figure 6). The sheath cells are fused into a syncytium, as are the socket cells. A third sensory neuron (SPC) is attached to the muscle at the base of the spicule and appears to signal the protracted or retracted state of the spicule to the spicule muscles. Spicule cells are born at the end of the L3 larval stage and differentiate and elongate during the second half of L4 (Sulston et al., 1980).
Spicule elongation and formation of the hardened, sclerotic cuticle that covers the spicule requires the outer socket cells but not the inner sheath cells or neurons (Jiang and Sternberg, 1999). Initially a spicule trace is formed in early L4 and elongates following anterior migrations of two B-lineage proctodeal cells (B.a(l/r)apapa and B.a(l/r)appv) that attach at the base of each spicule. Anterior migration of the proctodeal cells and elongation of the spicule require a signal of the TGFβ family (DBL-1 ligand, DAF-4 Type II receptor), probably generated by spicule muscles (Baird and Ellazar, 1999; Jiang and Sternberg, 1999). A signal of the FGF pathway (EGL-17 receptor) has also been implicated (Jiang and Sternberg, 1999). As the spicule cells migrate, they spread around a ridge of proctodeal cells which form a mold for the final, scalloped shape of the spicules (Sulston et al., 1980).
Cell fates in the complex B.a cell lineage that generates the cells of the spicules are influenced by a network of partially redundant intercellular signals coming from their neighbors in the rectal epithelium. Signals come from F and U on the anterior side and from Y.p on the posterior side as well as from cells of the B.a lineage itself (Chamberlin and Sternberg, 1993; Sulston et al., 1980). The F/U signal involves the lin-3/let-23 EGF pathway; lateral signaling involves the lin-12 pathway (Chamberlin and Sternberg, 1994).
The initial division of B.a is left/right (l/r) and this division generates equivalent cells (Figure 11B). These cells next divide along the anteroposterior (a/p) axis two times (Figure 11C, D). Unlike the initial l/r division, these a/p divisions generate daughter cells with different potentials, thereby giving rise to two identical l/r sets of four non-equivalent cells (8 cells in all). Next, the most anterior l/r pair migrates around the rectum to its ventral side (Figure 11D). These two cells and the most posterior l/r pair interdigitate at the midline respectively below and above the rectal channel, which thereby becomes surrounded by two rings of four cells (Figure 11E). As with some other bilateral pairs of cells that come together into a single longitudinal row (for example most of the Pn cells that generate ventral cord neurons), there is no correlation between left/right origin and final anteroposterior position in the row—the cells are equivalent initially and differences are induced by asymmetric signals after they arrive at the midline. Among the B-lineage cells, the difference in cell fates between the cells of the anterior ring and those of the posterior ring is induced by the lin-3/let-23 signal from F and U, which is modified by the further signals from Y.p and B.a cells. B.a cells also have further modulating influences on the primary signals (Figure 12; Chamberlin and Sternberg, 1994).
|
Figure 11. Patterning of cell fates in the B lineage. Left panels: Successive stages in the division of the B lineage, starting in the L1 larval stage. Arrows indicate the position of the cross section. In D, the B.a progeny cells have formed a ring around the rectal canal. In E, one cell has moved in front of another. At this stage, cell signaling specifies which cell will take the anterior fate and which the posterior fate (see text and Figure 12). Right panels: Nomarski photomicrographs showing the positions of cell nuclei around the rectal canal. Reprinted with permission from Chamberlin and Sternberg, 1993, Company of Biologists Ltd.
|
Figure 12. Multiple sets of interacting signals specify fates in the B lineage. (A) Positions of the cells at mid-L3 larval stage. Signals from F and U specify the fates of the anterior ring of cells. (B) Illustration of the multiple signals that specify the Bγ fate, which is assumed by B.alpp or B.arpp, depending on which of these two cells assumes the more anterior position. The primary signal is from F and U (1). Additional positional cues are provided by other B.a progeny (2) and Y.p (3). Lateral inhibitory cues reinforce the γ and δ decisions (4). Finally, cues from B.a progeny, which may be passive, prevent the F/U signal from acting inappropriately (5). Reprinted with permission from Chamberlin and Sternberg, 1993, Company of Biologists Ltd.
During the second half of postembryonic development, the posterior sensory structure common to both sexes, the phasmid, undergoes different development in the two sexes. The two phasmid socket cells, generated postembryonically from the T lineage, are added to the phasmid by L2. At this time, the two sexes are similar. In the adult hermaphrodite, the two sensory neurons are enveloped by a sheath cell and two layers of socket cells, the outermost, PHso2, attaching to the hypodermis. In the male, there is only a single socket cell, PHso2. PHso1 no longer envelops the sheath cell and develops some apparently neuronal characteristics, extending fingers into the sheath and having a basal body (Sulston et al., 1980). The functional significance of these changes is not known.
The proctodeum is the chamber just inside the cloaca where four channels come together: the vas deferens, the gut, and the two left/right spicule channels (Figure 13). The roof of the proctodeum is covered by sclerotic cuticle, forming a structure called the gubernaculum. The gubernaculum presumably guides the tips of the spicules down and out of the cloaca. The proctodeal chamber enlarges and the gubernaculum forms along with elongation of the spicules during the second half of L4. The gubernaculum is formed by B.paa with possible contributions from Bγl and Bγr (Sulston et al., 1980). Other cells making up the walls of the proctodeum include additional cells from the B.a and B.p lineages, K.a and K', and P12.pa (Figure 13).
The gut, which is of course attached to the cloacal canal from embryogenesis, joins the proctodeum via K.a and K'. The vas deferens attaches when the gonad arrives at the posterior during L4. At this time, K.a and K' as well as B.a(l/r)apaav extend out from the rectal wall and reach around the vas deferens cells, attaching to them. Attachment requires that the first-arriving cell of the gonad, the leader cell, be engulfed by either U.lp or U.rp. In the absence of U.lp and U.rp, K.a and K' show no affinity for the gonad and the connection is not made (Sulston et al., 1980).
Differentiation of the proctodeum is affected by many of the genes required for patterning the fates and progeny of B (see above; section 4.4; Chamberlin et al., 1999; Sewell et al., 2003). Attachment of the proctodeal wall to the gonad requires the DM domain transcription factor mab-23, which is expressed in the attaching cells K.a and K' and in cells of the U lineage, including the ones necessary to engulf the leader cell. In a mab-23 mutant, rectal cells fail to extend towards and contact the gonadal cells, which apparently remain unattached, allowing sperm to pass into the coelomic cavity (Lints and Emmons, 2002).
Male behavior is augmented extensively over that of the hermaphrodite to bring about transfer of the gametes. If a male contacts a hermaphrodite, he makes attempts at copulation. When hermaphrodites are present, males remain with them, but if hermaphrodites are absent, males engage in mate-searching (Lipton et al., 2004; see Male mating behavior). The male nervous system has almost the same set of neurons as the hermaphrodite plus some 79 additional neurons, almost all in the tail. Some neurons in the common set are modified in the male. For example, PHC, which has a process in the tail spike, has a striated rootlet in the male and many additional synapses over that of the hermaphrodite (Sulston et al., 1980). Similarly, PDB, an interneuron, has many more branches in the male pre-anal ganglion than in the hermaphrodite. How extensive such sex-specific modification of common neurons is will remain unknown until there is a reconstruction of the male nervous system.
The neurons in the male tail that are born during postembryonic development are the last neurons in the nervous system to differentiate. Some of these differentiate in a matter of hours at the end of the L4 larval stage, sending out their axons into the existing neuropil to find their targets.
The muscles generated during embryogenesis are the same in males and hermaphrodites. Additional muscles arise postembryonically from a single mesoblast, the M cell, the initial divisions of which during L1 are also similar in the two sexes and generate 14 new body wall muscles (Sulston and Horvitz, 1977). However, whereas in the hermaphrodite two M cell progeny divide further to make sex muscles, in the male these same cells plus four additional cells that in the hermaphrodite become coelomocytes are also sex mesoblasts. In the hermaphrodite, the two sex mesoblasts migrate anteriorly and generate vulval muscles, while in the male the six sex mesoblasts migrate posteriorly into the tail.
The sex mesoblasts (three left-right symmetrical pairs) begin to divide again during the L3 larval stage and by the end of L3 have generated 41 new muscles and a coelomocyte (Sulston et al., 1980). Ten of the new muscles run longitudinally along the body wall, apparently supplementing the embryonic body wall muscles in this region. Fifteen form muscles called diagonal muscles that attach between the ventral and mid-lateral body wall and aid in the curling of the tail during copulation (Loer and Kenyon, 1993). The remaining 16 cells differentiate into muscles that control the spicules (8) and gubernaculum (4), or that run across the tail posterior of the anus to pull the dorsal and ventral body walls together (4).
In addition to having these new muscles, the male enlarges the intestinal sphincter, which surrounds the gut and controls its opening into the proctodeal chamber, and reorganizes the orientation and attachments of the myofilaments in the anal depressor muscle (Sulston et al., 1980). Each of these changes presumably aid ejaculation.
The adult male gonad, radically distinct from that of the hermaphrodite, develops from a superficially identical gonad primordium located at the midbody. The gonadal primordium consists of two germ cells (Z2, Z3) flanked by two somatic precursor cells (Z1, Z4). In both sexes, the developing gonad grows out from this primordium to establish its elongated, tubular form. Compared to that of the hermaphrodite, outgrowth in the male occurs in the opposite orientation with respect to the ultimate polarity of the gonad. That is, whereas in the hermaphrodite the midbody location of the gonad primordium will become the proximal end of the gonad (the site of attachment of the uterus to the vulval opening) and the developing gonad will grow out towards the distal end of the ovary (the site of mitotically dividing germ cells), in the male the site of the primordium becomes the distal end of the testes and the developing gonad grows out towards the future site of attachment and opening of the vas deferens at the cloaca in the tail (Kimble and Hirsh, 1979). Outgrowth in the male is lead by a linker cell instead of by a distal tip cell as in the hermaphrodite. The linker cell has to be engulfed upon arriving at the cloaca by rectal epithelial cells of the U lineage before additional rectal cells (K.a, K', and B.a(l/r)apaav) attach to the vas deferens (Sulston et al., 1980).
A second difference between male and hermaphrodite gonads is that, whereas the hermaphrodite gonad is two-armed and symmetric, that of the male gonad is one-armed and asymmetric. Development of asymmetry in the male gonad appears already after the second division of Z1 and Z4, when Z4.a migrates anteriorly to join Z1.p, and Z1.a migrates posteriorly to join Z4.p. Z1.a and Z4.p will not divide again and will remain at midbody, becoming the distal tip cells at the distal end of the testes. These, like their hermaphrodite counterparts, are required to maintain distal germ cells in mitosis. Interactions among descendants of Z1.p and Z4.a result in the selection of a single cell to function as the linker cell. This is analogous to the cell-cell interactions between Z1.p and Z4.a descendants (Z1.ppp and Z4.aaa) in the hermaphrodite that select one cell to assume the anchor cell fate (Kimble, 1981). Anchor cell and male linker cell have the similar function of attaching the proximal end of the gonad to the opening to the exterior. Thus, in spite of radically differing organization, underlying analogies in cell fate assignments in the male and hermaphrodite lineages are clearly discernable (Kimble and Hirsh, 1979). Both sexes appear to utilize a single underlying gonad developmental program that is modulated at points to suit the sex of the animal. Not surprisingly, a number of developmental genes, including lin-12 and the Wnt receptor lin-17, are required for gonad development in both sexes. One transcription factor that appears to play a more significant role in specifying the male pathway is the forkhead transcription factor gene fkh-6 (Chang et al., 2004).
I thank K. Chow, R. Ghosh, G. Kleemann, R. Lints, and P. Sternberg for their comments on the manuscript and D. Portman for contribution to Figure 6. Work in the author's laboratory is funded by the US National Institutes of Health. SWE is the Siegfried Ullmann Professor of Molecular Genetics.
Austin, J., and Kenyon, C. (1994). Cell contact regulates neuroblast formation in the Caenorhabditis elegans lateral epidermis. Development 120, 313–323. Abstract
Baird, S.E., and Ellazar, S.A. (1999). TGFbeta-like signaling and spicule development in Caenorhabditis elegans. Dev. Biol. 212, 93–100. Abstract Article
Baird, S.E., and Emmons, S.W. (1990). Properties of a class of genes required for ray morphogenesis in Caenorhabditis elegans. Genetics 126, 335–344. Abstract
Baird, S.E., Fitch, D.H., Kassem, I.A., and Emmons, S.W. (1991). Pattern formation in the nematode epidermis: determination of the arrangement of peripheral sense organs in the C. elegans male tail. Development 113, 515–526. Abstract
Barr, M.M., DeModena, J., Braun, D., Nguyen, C.Q., Hall, D.H., and Sternberg, P.W. (2001). The Caenorhabditis elegans autosomal dominant polycystic kidney disease gene homologs lov-1 and pkd-2 act in the same pathway. Curr. Biol. 11, 1341–1346. Abstract Article
Barr, M.M., and Sternberg, P.W. (1999). A polycystic kidney-disease gene homologue required for male mating behaviour in C. elegans. Nature 401, 386–389. Abstract Article
Chamberlin, H.M., Brown, K.B., Sternberg, P.W., and Thomas, J.H. (1999). Characterization of seven genes affecting Caenorhabditis elegans hindgut development. Genetics 153, 731–742. Abstract
Chamberlin, H.M., Palmer, R.E., Newman, A.P., Sternberg, P.W., Baillie, D. L., and Thomas, J.H. (1997). The PAX gene egl-38 mediates developmental patterning in Caenorhabditis elegans. Development 124, 3919–3928. Abstract
Chamberlin, H.M., and Sternberg, P.W. (1993). Multiple cell interactions are required for fate specification during male spicule development in Caenorhabditis elegans. Development 118, 297–324. Abstract
Chamberlin, H.M., and Sternberg, P.W. (1994). The lin-3/let-23 pathway mediates inductive signalling during male spicule development in Caenorhabditis elegans. Development 120, 2713–2721. Abstract
Chamberlin, H.M., and Sternberg, P.W. (1995). Mutations in the Caenorhabditis elegans gene vab-3 reveal distinct roles in fate specification and unequal cytokinesis in an asymmetric cell division. Dev. Biol. 170, 679–689. Abstract Article
Chang, W., Tilman, C., Thoemke, K., Markussen, F.H., Mathies, L.D., Kimble, J., and Zarkower, D. (2004). A forkhead protein controls sexual identity of the C. elegans male somatic gonad. Development 131, 1425–1436. Abstract Article
Chin-Sang, I.D., Moseley, S.L., Ding, M., Harrington, R.J., George, S.E., and Chisholm, A.D. (2002). The divergent C. elegans ephrin EFN-4 functions inembryonic morphogenesis in a pathway independent of the VAB-1 Eph receptor. Development 129, 5499–5510. Abstract Article
Chisholm, A. (1991). Control of cell fate in the tail region of C. elegans by the gene egl-5 . Development 111, 921–932. Abstract
Chisholm, A.D., and Hodgkin, J. (1989). The mab-9 gene controls the fate of B, the major male-specific blast cell in the tail region of Caenorhabditis elegans. Genes Dev. 3, 1413–1423. Abstract
Chow, K.L., and Emmons, S.W. (1994). HOM-C/Hox genes and four interacting loci determine the morphogenetic properties of single cells in the nematode male tail. Development 120, 2579–2592. Abstract
Chow, K.L., Hall, D.H., and Emmons, S.W. (1995). The mab-21 gene of Caenorhabditis elegans encodes a novel protein required for choice of alternate cell fates. Development 121, 3615–3626. Abstract
Cowing, D.W., and Kenyon, C. (1992). Expression of the homeotic gene mab-5 during Caenorhabditis elegans embryogenesis. Development 116, 481–490. Abstract
Dalpe, G., Zhang, L.W., Zheng, H., and Culotti, J.G. (2004). Conversion of cell movement responses to Semaphorin-1 and Plexin-1 from attraction to repulsion by lowered levels of specific RAC GTPases in C. elegans. Development 131, 2073–2088. Abstract Article
Emmons, S.W., and Sternberg, P.W. (1997). Male development and mating behavior. In: C. elegans II. D.L. Riddle, T. Blumenthal, B.J. Meyer, and J.R. Priess, eds. (Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press), pp. 295–334.
Ferreira, H.B., Zhang, Y., Zhao, C., and Emmons, S.W. (1999). Patterning of Caenorhabditis elegans posterior structures by the Abdominal-B homolog, egl-5. Dev. Biol. 207, 215–228. Abstract Article
Fitch, D.H., Emmons, .S.W. (1995). Variable cell positions and cell contacts underlie morphological evolution of the rays in the male tails of nematodes related to Caenorhabditis elegans. Dev. Biol. 170(2), 564–582. Article
Fujii, T., Nakao, F., Shibata, Y., Shioi, G., Kodama, E., Fujisawa, H., and Takagi, S. (2002). Caenorhabditis elegans PlexinA, PLX-1, interacts with transmembrane semaphorins and regulates epidermal morphogenesis. Development 129, 2053–2063. Abstract
Ginzburg, V.E., Roy, P.J., and Culotti, J.G. (2002). Semaphorin 1a and semaphorin 1b are required for correct epidermal cell positioning and adhesion during morphogenesis in C. elegans. Development 129, 2065–2078. Abstract
Greenwald, I.S., Sternberg, P.W., and Horvitz, H.R. (1983). The lin-12 locus specifies cell fates in Caenorhabditis elegans. Cell 34, 435–444. Abstract Article
Hahn, A.C., and Emmons, S.W. (2003). The roles of an ephrin and a semaphorin in patterning cell-cell contacts in C. elegans sensory organ development. Dev. Biol. 256, 379–388. Abstract Article
Herman, M.A., and Horvitz, H.R. (1994). The Caenorhabditis elegans gene lin-44 controls the polarity of asymmetric cell divisions. Development 120, 1035–1047. Abstract
Herman, M.A., Vassilieva, L.L., Horvitz, H.R., Shaw, J.E., and Herman, R.K. (1995). The C. elegans gene lin-44, which controls the polarity of certain asymmetric cell divisions, encodes a Wnt protein and acts cell nonautonomously. Cell 83, 101–110. Abstract Article
Hill, K.L., Harfe, B.D., Dobbins, C., A., and L'Hernault, S.W. (2000). dpy-18 encodes an alpha subunit of prolyl-4-hydroxylase in Caenorhabditis elegans. Genetics 155, 1139–1148.
Hodgkin, J. (1983). Male phenotypes and mating efficiency in Caenorhabditis elegans. Genetics 103, 43–64.
Hunter, C.P., Harris, J.M., Maloof, J.N., and Kenyon, C. (1999). Hox gene expression in a single Caenorhabditis elegans cell is regulated by a caudal homolog and intercellular signals that inhibit wnt signaling. Development 126, 805–814. Abstract
Ikegami, R., Zheng, H., Ong, S.H., and Culotti, J. (2004). Integration of semapho rin-2 A/MAB-20, ephrin-4, and UNC-129 TGF-beta signaling pathways regulates sorting of distinct sensory rays in C. elegans. Dev. Cell 6, 383–395. Abstract Article
Jiang, L.I., and Sternberg, P.W. (1998). Interactions of EGF, Wnt and HOM-C genes specify the P12 neuroectoblast fate in C. elegans. Development 125, 2337–2347. Abstract
Jiang, L.I., and Sternberg, P.W. (1999). Socket cells mediate spicule morphogenesis in Caenorhabditis elegans males. Dev. Biol. 211, 88–99. Abstract Article
Jiang, M., Ryu, J., Kiraly, M., Duke, K., Reinke, V., and Kim, S.K. (2001). Genome-wide analysis of developmental and sex-regulated gene expression profiles in Caenorhabditis elegans. Proc. Natl. Acad. Sci. USA 98, 218–223. Abstract Article
Johnson, A.D., Fitzsimmons, D., Hagman, J., and Chamberlin, H.M. (2001). EGL-38 Pax regulates the ovo-related gene lin-48 during Caenorhabditis elegans organ development. Development 128, 2857–2865. Abstract
Kenyon, C. (1986). A gene involved in the development of the posterior body region of C. elegans. Cell 46, 477–487. Abstract Article
Kimble, J. (1981). Alterations in cell lineage following laser ablations of cells in the somatic gonad of Caenorhabditis elegans. Dev. Biol. 87, 286–300. Abstract Article
Kimble, J., and Hirsh, D. (1979). The postembryonic cell lineages of the hermaphrodite and male gonads in Caenorhabditis elegans. Dev. Biol. 70, 396–417. Abstract Article
Ko, F.C., and Chow, K.L. (2002). A novel thioredoxin-like protein encoded by the C. elegans dpy-11 gene is required for body and sensory organ morphogenesis. Development 129, 1185–1194. Abstract
Liang, J., Lints, R., Foehr, M.L., Tokarz, R., Yu, L., Emmons, S.W., Liu, J., and Savage-Dunn, C. (2003). The Caenorhabditis elegans schnurri homolog sma-9 mediates stage- and cell type-specific responses to DBL-1 BMP-related signaling. Development 130, 6453–6464. Abstract Article
Lints, R., and Emmons, S.W. (1999). Patterning of dopaminergic neurotransmitter identity among Caenorhabditis elegans ray sensory neurons by a TGFbeta family signaling pathway and a Hox gene. Development 126, 5819–5831. Abstract
Lints, R., and Emmons, S.W. (2002). Regulation of sex-specific differentiation and mating behavior in C. elegans by a new member of the DM domain transcription factor family. Genes Dev. 16, 2390–2402. Abstract Article
Lints, R., Jia, L., Kim, K., Li, C., and Emmons, S.W. (2004). Axial patterning of C. elegans male sensilla identities by selector genes. Dev. Biol. 269, 137–151. Abstract Article
Lipton, J., Kleemann, G., Ghosh, R., Lints, R., and Emmons, S.W. (2004). Mate searching in Caenorhabditis elegans: a genetic model for sex drive in a simple invertebrate. J. Neurosci. 24, 7427–7434. Abstract Article
Liu, K.S., and Sternberg, P.W. (1995). Sensory regulation of male mating behavior in Caenorhabditis elegans. Neuron 14, 79–89. Abstract Article
Loer, C.M., and Kenyon, C.J. (1993). Serotonin-deficient mutants and male mating behavior in the nematode Caenorhabditis elegans. J. Neurosci. 13, 5407–5417. Abstract
Maloof, J.N., Whangbo, J., Harris, J.M., Jongeward, G.D., and Kenyon, C. (1999). A Wnt signaling pathway controls hox gene expression and neuroblast migration in C. elegans. Development 126, 37–49. Abstract
Morita, K., Chow, K.L., and Ueno, N. (1999). Regulation of body length and male tail ray pattern formation of Caenorhabditis elegans by a member of TGF-beta family. Development 126, 1337–1347. Abstract
Nguyen, C.Q., Hall, D.H., Yang, Y., and Fitch, D.H. (1999). Morphogenesis of the Caenorhabditis elegans male tail tip. Dev. Biol. 207, 86–106. Abstract Article
Portman, D.S., and Emmons, S.W. (2000). The basic helix-loop-helix transcription factors LIN-32 and HLH-2 function together in multiple steps of a C. elegans neuronal sublineage. Development 127, 5415–5426. Abstract
Portman, D.S., and Emmons, S.W. (2004). Identification of C. elegans sensory ray genes using whole-genome expression profiling. Dev. Biol. 270, 499–512. Abstract Article
Roy, P.J., Zheng, H., Warren, C.E., and Culotti, J.G. (2000). mab-20 encodes Semaphorin-2a and is required to prevent ectopic cell contacts during epidermal morphogenesis in Caenorhabditis elegans. Development 127, 755–767. Abstract
Salser, S.J., and Kenyon, C. (1996). A C. elegans Hox gene switches on, off, on and off again to regulate proliferation, differentiation and morphogenesis. Development 122, 1651–1661. Abstract
Salser, S.J., Loer, C.M., and Kenyon, C. (1993). Multiple HOM-C gene interactions specify cell fates in the nematode central nervous system. Genes Dev. 7, 1714–1724. Abstract
Savage-Dunn, C., Das, P., Finelli, A.L., Townsend, S.R., Sun, C.Y., Baird, S. E., and Padgett, R.W. (1996). Caenorhabditis elegans genes sma-2, sma-3 and sma-4 define a conserved family of transforming growth factor beta pathway components. Proc. Natl. Acad. Sci. USA 93, 790–794. Abstract Article
Schinkmann, K., and Li, C. (1992). Localization of FMRFamide-like peptides in Caenorhabditis elegans. J. Comput. Neurol. 316, 251–260. Article
Sewell, S.T., Zhang, G., Uttam, A., and Chamberlin, H.M. (2003). Developmental patterning in the Caenorhabditis elegans hindgut. Dev. Biol. 262, 88–93. Abstract Article
Shen, M.M., and Hodgkin, J. (1988). mab-3, a gene required for sex-specific yolk protein expression and a male-specific lineage in C. elegans. Cell 54, 1019–1031. Abstract Article
Sternberg, P.W., and Horvitz, H.R. (1988). lin-17 mutations of Caenorhabditis elegans disrupt certain asymmetric cell divisions. Dev. Biol. 130, 67–73. Abstract Article
Sulston, J.E., Albertson, D.G., and Thomson, J.N. (1980). The Caenorhabditis elegans male: postembryonic development of nongonadal structures. Dev. Biol. 78, 542–576. Abstract Article
Sulston, J.E., and Horvitz, H.R. (1977). Post-embryonic cell lineages of the nematode, Caenorhabditis elegans. Dev. Biol. 56, 110–156. Abstract Article
Sulston, J.E., and White, J.G. (1980). Regulation and cell autonomy during postembryonic development of Caenorhabditis elegans. Dev. Biol. 78, 577–597. Abstract Article
Sutherlin, M.E., and Emmons, S.W. (1994). Selective lineage specification by mab-19 during Caenorhabditis elegans male peripheral sense organ development. Genetics 138, 675–688. Abstract
Suzuki, Y., Yandell, M.D., Roy, P.J., Krishna, S., Savage-Dunn, C., Ross, R. M., Padgett, R.W., and Wood, W.B. (1999). A BMP homolog acts as a dose-dependent regulator of body size and male tail patterning in Caenorhabditis elegans. Development 126, 241–250. Abstract
Teng, Y., Girard, L., Ferreira, H.B., Sternberg, P.W., and Emmons, S.W. (2004). Dissection of cis-regulatory elements in the C. elegans Hox gene egl-5 promoter. Dev. Biol. 276, 476–492. Abstract Article
Troemel, E.R., Chou, J.H., Dwyer, N.D., Colbert, H.A., and Bargmann, C.I. (1995). Divergent seven transmembrane receptors are candidate chemosensory receptors in C. elegans. Cell 83, 207–218. Abstract Article
Waring, D.A., and Kenyon, C. (1990). Selective silencing of cell communication influences anteroposterior pattern formation in C. elegans. Cell 60, 123–131. Abstract Article
Winter, A.D., and Page, A.P. (2000). Prolyl 4-hydroxylase is an essential procollagen-modifying enzyme required for exoskeleton formation and maintenance of body shape in the nematode Caenorhabditis elegans. Mol. Cell. Biol. 20, 4084–4093. Article
Woollard, A., and Hodgkin, J. (2000). The Caenorhabditis elegans fate-determining gene mab-9 encodes a T-box protein required to pattern the posterior hindgut. Genes Dev. 14, 596–603. Abstract
Wrischnik, L.A., and Kenyon, C.J. (1997). The role of lin-22, a hairy/enhancer of split homolog, in patterning the peripheral nervous system of C. elegans. Development 124, 2875–2888. Abstract
Yi, W., Ross, J.M., and Zarkower, D. (2000). mab-3 is a direct tra-1 target gene regulating diverse aspects of C. elegans male sexual development and behavior. Development 127, 4469–4480. Abstract
Yu, H., Pretot, R.F., Burglin, T.R., and Sternberg, P.W. (2003). Distinct roles of transcription factors EGL-46 and DAF-19 in specifying the functionality of a polycystin-expressing sensory neuron necessary for C. elegans male vulva location behavior. Development and Psychopathology 130, 5217–5227.
Yu, R.Y., Nguyen, C.Q., Hall, D.H., and Chow, K.L. (2000). Expression of ram-5 in the structural cell is required for sensory ray morphogenesis in Caenorhabditis elegans male tail. EMBO J. 19, 3542–3555. Abstract Article
Zhang, Y., and Emmons, S.W. (1995). Specification of sense-organ identity by a Caenorhabditis elegans Pax-6 homologue. Nature 377, 55–59. Abstract Article
Zhao, C., and Emmons, S.W. (1995). A transcription factor controlling development of peripheral sense organs in C. elegans. Nature 373, 74–78. Abstract Article
Zhao, X., Yang, Y., Fitch, D.H., and Herman, M.A. (2002). TLP-1 is an asymmetric cell fate determinant that responds to Wnt signals and controls male tail tip morphogenesis in C. elegans. Development 129, 1497–1508. Abstract
*Edited by Barbara J. Meyer. Last revised October 20, 2005. Published November 10, 2005. This chapter should be cited as: Emmons, S.W. Male development (November 10, 2005), WormBook, ed. The C. elegans Research Community, WormBook, doi/10.1895/wormbook.1.33.1, http://www.wormbook.org.
Copyright: © 2005 Scott W. Emmons. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
§To whom correspondence should be addressed. E-mail: [email protected]
 All WormBook content, except where otherwise noted, is licensed under a Creative Commons Attribution License.
All WormBook content, except where otherwise noted, is licensed under a Creative Commons Attribution License.