Embryo series courtesy of Einhard Schierenberg
Embryo series courtesy of Einhard SchierenbergTable of Contents
Abstract
Oscheius tipulae is a common soil nematode of the same family as C. elegans (Rhabditidae), which presents the same hermaphroditic mode of reproduction and is easily cultured in the same conditions. Oscheius tipulae has been used as a developmental genetic model system to study vulva formation. Compared to C. elegans, it has a simpler vulval cell lineage, a reduced competence group and a different mechanism of vulval cell fate patterning. The spectrum of vulval phenotypes obtained in genetic screens differs from that found in C. elegans. Its easy isolation from soil and the availability of numerous wild isolates of O. tipulae from all over the world facilitate population genetic and microevolutionary studies, especially of the evolution of cell lineage. The Oscheius genus also presents many species with interesting evolutionary changes in mode of reproduction, gonad development, body size, etc.
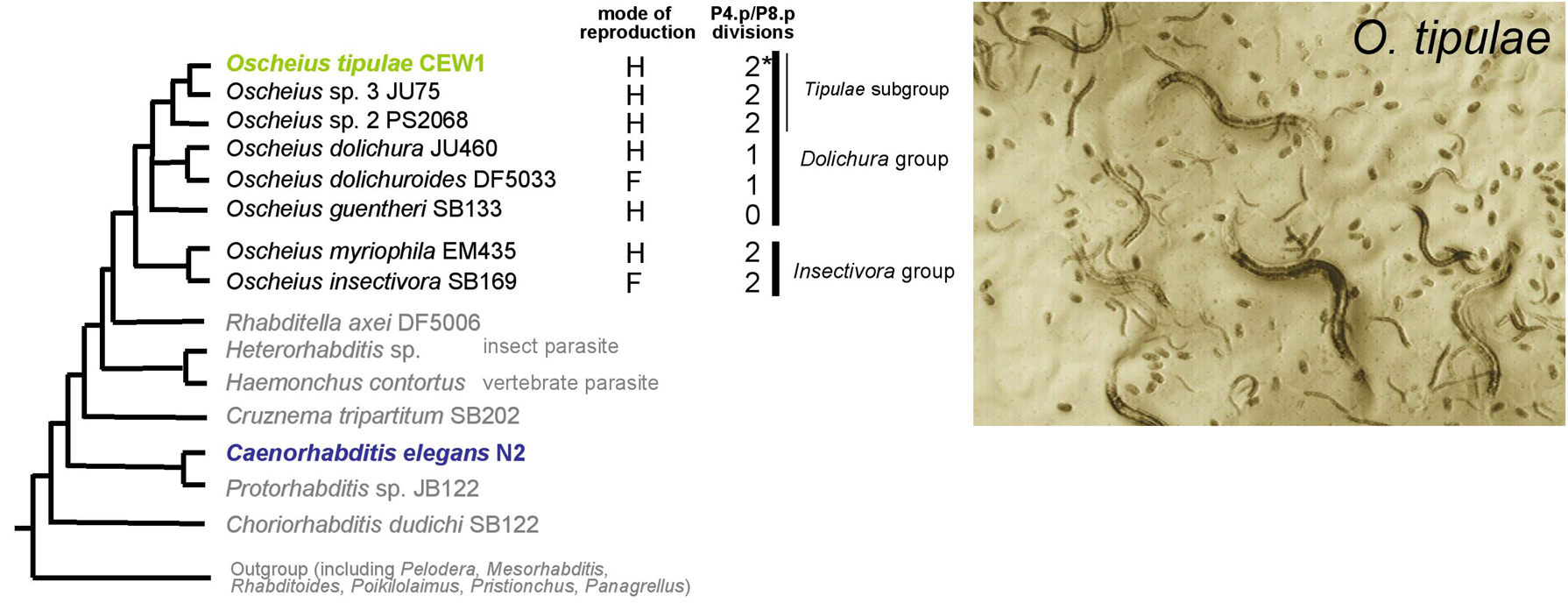
Oscheius tipulae is a nematode of the same family as C. elegans (Rhabditidae), commonly found in soil. Its relationship to C. elegans is more distant than that of members of the Caenorhabditis genus, yet it is more closely related to C. elegans than Pristionchus pacificus and Brugia malayi are (Figure 1). Compared to C. elegans, Oscheius tipulae shares a common ancestor with the insect parasite Heterorhabditis and the vertebrate parasites Strongylida (Blaxter et al., 1998; see The phylogenetic relationships of Caenorhabditis and other rhabditids).
|
Figure 1. Phylogenetic relationships of some nematodes, showing the position of Oscheius tipulae. After Blaxter et al. (1998), Félix et al. (2001), The phylogenetic relationships of Caenorhabditis and other rhabditids. Branch lengths have no meaning. Modes of reproduction: H, hermaphrodites and facultative males; F, females and males. Some species in the Dolichura subgroup produce F1 hybrids, some of them appearing somewhat fertile (M.-A. F., unpublished). Number of division rounds of P4.p/P8.p : *indicates a polymorphism between 0 and 2 division rounds (Delattre and Félix, 2001a).
The Oscheius genus comprises two main subclades (Dolichura and Insectivora groups; see The phylogenetic relationships of Caenorhabditis and other rhabditids; Sudhaus, 1976; Sudhaus and Hooper, 1994; Figure 1). One distinctive character between these two clades appears to be the smaller size of the animals in the Dolichura clade (that includes Oscheius tipulae) (Flemming et al., 2000) (M.-A. F., unpublished). The Dolichura group comprises Oscheius guentheri, which is a species with a reduced posterior-gonadal arm (Sudhaus and Hooper, 1994), plus two subgroups, of which two representative species are Oscheius tipulae and O. dolichura, respectively (see The phylogenetic relationships of Caenorhabditis and other rhabditids). Historically, Oscheius dolichura (Schneider, 1866) was one of the first nematode species (before C. elegans) described to have an androdioceous mode of reproduction (Maupas, 1900; see English translation: http://www.wormbase.org/papers/1900-maupas/index.html).
Oscheius tipulae was first described by Lam and Webster (Lam and Webster, 1971). The species was brought to persisting live culture (strain SB128) and redescribed by Sudhaus (1993). It was called O. tipulae because these researchers found it associated with the larvae of the insect Tipula paludosa (a dipteran). The CEW1 strain was found by C.E. Winter in soil in São Paulo, Brazil around 1992 (Winter, 1992). Because of taxonomic confusions, the CEW1 strain has been variably named Dolichorhabditis sp. or D. brevesophaga or Oscheius sp. 1 before it was decided to be an isolate of Oscheius tipulae (Dichtel-Danjoy and Félix, 2004), which is the valid name.
O. tipulae and its two closely related species, provisionally called Oscheius spp. 2 and 3 (Figure 1), are hard to distinguish morphologically, but can be distinguished through mating experiments and molecular features (Félix et al., 2001).
Oscheius tipulae is one of the most common nematode species in soil of different (non-desert) parts of the world. It is both widespread and abundant in soil samples. It is mostly (or exclusively) found in the dauer stage (like C. elegans), raising the question of whether it proliferates in soil. It was originally isolated from Tipula larvae, but this may not be a particularly specific association. O. tipulae is more heat-resistant in the lab than O. spp. 2 and 3 (Félix et al., 2001) and has been found in tropical as well as temperate regions.
Oscheius tipulae is easy to isolate (see Isolation of C. elegans and related nematodes). Species of the Oscheius genus are characterized in particular by their long rectum (see Figure 5 in Isolation of C. elegans and related nematodes).
Available wild isolates can be found at the CGC (http://elegans.swmed.edu/CGC/), the NYU Rhabditidae collection (http://www.nyu.edu/projects/fitch/WSRN/) or in the Félix lab collection (http://www2.ijm.jussieu.fr/worms).
AFLP (Amplified Fragment Length Polymorphism) analysis on 53 wild isolates from all over the world suggest a genetic diversity at least three-fold higher than that measured with the same method in C. elegans (Barrière and Félix, 2005), with no genetic differentiation between continents, yet a clear structure at a smaller scale between three local populations sampled in France (100 km scale), and a relatively high local diversity (D. Baïlle and M.-A. F., unpublished).
Oscheius tipulae has the same mode of reproduction as C. elegans (self-fertile hermaphrodites and facultative males), and can be cultured and frozen in the same conditions. The lifecycle of the reference strain CEW1 lasts approximately 3 days at 25°C, 4 days at 23°C and 5–6 days at 20°C. Oscheius tipulae develops through four juvenile stages and an alternative dauer stage like C. elegans. The animals are smaller and slower than C. elegans. Eggs are laid at the one-cell stage. Adult hermaphrodites rarely bag upon starvation. The larvae are clear in Nomarski optics. The gonad is syncytial with a central medial region where injected material can diffuse (rachis).
Oscheius tipulae reproduces through XX hermaphrodites and XO males like C. elegans.
The X chromosome non-disjunction rate of the reference strain CEW1 is lower than that of C. elegans N2 and males are not easy to induce by heat-shock. However, other O. tipulae strains such as PS959 have a higher proportion of males. Males reproduce less well at 25°C and above than at lower temperatures; therefore, crossing plates are better maintained at 23°C (or below).
Mutagenesis and genetic analysis are basically performed as in C. elegans (Brenner, 1974). Because the germ line divides later than that of C. elegans, mutagenesis is best performed on young adults (rather than L4-stage animals). The concentration of EMS used for C. elegans (50 mM) is not as efficient nor as lethal as in C. elegans; therefore, mutageneses were performed using an EMS concentration of 100–150 mM, or using TMP-UV irradiation.
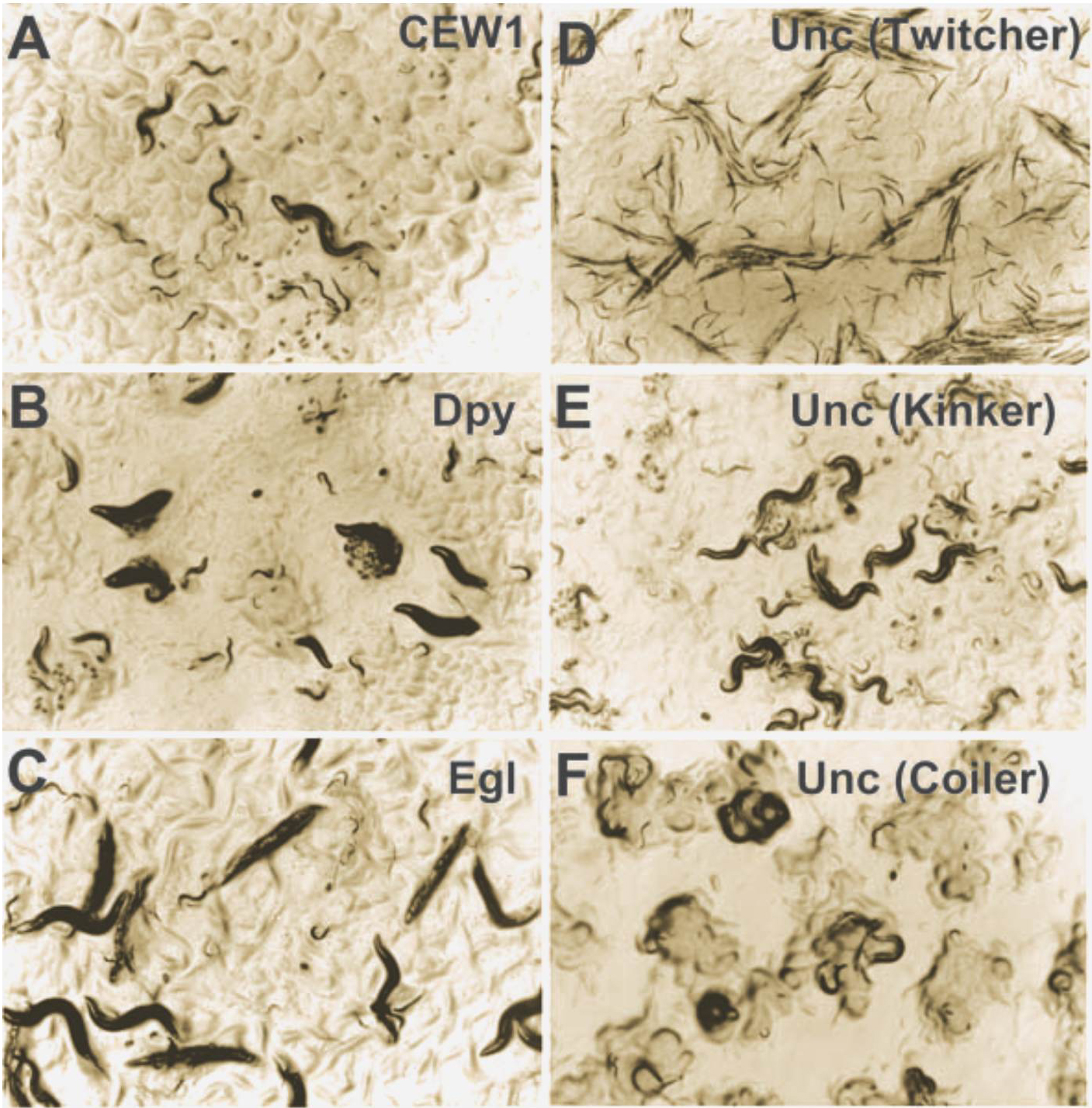
Screens for visible mutants and for mutants with a defective vulva (see below, Section 6.) have been performed. 79 visible markers were isolated: 24 with a Dumpy or Small phenotype, 5 Rollers, 2 Longs, 1 Constipated and 47 Uncs (twitchers, kinkers, sluggish worms, coilers including one dominant allele; Figure 2). Slightly surprisingly, 35 out of 74 mutants are X-linked (especially 26/43 Uncs compared to 3/23 Dpys, M.-L. Dichtel and M.-A. F., unpublished). Mutants with a Him phenotype (recessive or dominant) were also isolated, as well as one mutant with a partially penetrant left-right inversion of the body (cov-7; Delattre and Félix, 2001c; Louvet-Vallée et al., 2003).
|
Figure 2. Wild type and some mutants of Oscheius tipulae. A. Wild type CEW1. An adult hermaphrodite, embryos and various larval stages are visible. B. Dumpy phenotype (mutant sy536). C. Egg-laying defective phenotype (mutant sy537) with bags of larvae. D. Twitcher phenotype (mutant sy486). E. Kinker phenotype (mutant sy474). F. Coiler phenotype (mutant sy445).
The apparent haploid number of chromosomes observed by DAPI staining of the germ line is six, as in C. elegans. G. Maro, M.-L. Dichtel and M.-A. Félix defined (by genetic complementation or linkage data; unpublished) 6 dpy loci, 7 unc loci and 2 lon loci. The unc, dpy, and lon loci are provisionally designated by a letter to avoid confusion with C. elegans loci. Rough linkage data gave six linkage groups, which may or may not cover the six chromosomes:
| 5 autosomal linkage groups: |
| A. unc-a(sy486) (Twitcher), unc-c(mf43), unc-d(sy457), unc-e(mf42), dpy-a(mf1), dpy-e(sy517), iov-1(mf86) (not ordered) |
| B. dpy-c(sy471), lon-a(sy475) |
| C. unc-b(mf29) |
| D. dpy-f(sy518) |
| E. con-a(sy462) |
| X-linked: |
| unc-f(sy474), unc-g(sy485), rol-a(sy472), lon-b(sy502) (not ordered) |
A few mutants are at CGC. Others can be found at http://www2.ijm.jussieu.fr/worms. The gene nomenclature is temporary, since these loci are not homologs to the corresponding C. elegans genes. Once their molecular identity is determined, the correct nomenclature is, for example, Oti-lin-39 for the Oscheius tipulae lin-39 ortholog.
Injection of morpholinos results in a partial inactivation of the corresponding gene product, for example Oti-lin-39 (Louvet-Vallée et al., 2003) or Oti-let-60 (Dichtel-Danjoy and Félix, 2004). However, as in many non-Caenorhabditis nematodes, RNA interference does not appear to work efficiently: injection of double-stranded RNA for various genes has not produced any phenotype.
DNA-mediated transformation has been unsuccessful so far, but recent studies in Strongyloides stercoralis suggest that using a 3′UTR from the species may be crucial for transgene expression (Li et al., 2006). This remains to be tested in Oscheius tipulae. Plasmids injected into the mother's female germ line have been detected by PCR in the F1 progeny (M.-L. Dichtel, unpublished).
From rough estimates based on hybridization of a genomic library, the genome size of Oscheius tipulae appears similar to that of C. elegans (M. Delattre, unpublished). Its GC content was determined to be close to 43% (Ahn and Winter, 2005). Codon biases of highly expressed genes such as vitellogenins appear slightly different from those in C. elegans (Winter et al., 1996). Transcription in operons also occurs in O. tipulae as in C. elegans, with SL1 and SL2 trans-splicing; these SL sequences are similar yet slightly different from those of C. elegans (Evans et al., 1997).
Libraries from Oscheius tipulae CEW1 include a cDNA library in λZAP II described in Félix et al. (2000) and a genomic library in λFIX II described in Louvet-Vallée et al. (2003) (available on request: [email protected]).
Oscheius tipulae taxon number in sequence databases is 141969; because of changes in the species’ name that have been associated with particular strains in the literature (see Section 1.2), some of the sequences may not appear in a direct search for “Oscheius tipulae” in databases. In addition, a few dozen ESTs have been sequenced (M.-A. F. and P.W. Sternberg, unpublished).
Gonad development and morphology of O. tipulae are overall similar to those of C. elegans, except that germ line divisions are delayed. However, O. guentheri presents an interesting case of variable partial reduction of the posterior gonadal arm, which appears to result from a lesser activity of the posterior distal tip cell in its roles in arm migration and germ line proliferation (Félix and Sternberg, 1996).
Anchor cell determination is stochastic and unbiased in O. tipulae as in C. elegans, but biased towards Z4.aaa becoming the anchor cell in Oscheius guentheri (Félix and Sternberg, 1996; Sudhaus and Hooper, 1994).
Oscheius tipulae has been mostly studied for its vulva development (see Evolution of development in nematodes related to C. elegans). Several features make it a particularly interesting comparison to C. elegans. Its vulval cell lineage is different, and simpler than that of C. elegans. In particular, it is much easier to distinguish the 1° and 2° precursor cell fates by the number of division rounds they undergo (three and two, respectively) (Sommer and Sternberg, 1995; Figure 3). The vulval competence group is composed of P(4-8).p: P3.p is not competent (unlike in C. elegans). The adult vulva is formed of six superposed rings, instead of seven in C. elegans (Louvet-Vallée et al., 2003). Most strikingly, anchor cell ablations demonstrate that the anchor cell is required after P6.p division for its daughters to acquire a 1° fate: if the anchor cell is ablated in the mid-L3 stage, P(5-7).p all adopt a 2° lineage pattern. The same spatial pattern of fates is thus induced by different mechanisms in O. tipulae and C. elegans (Félix and Sternberg, 1997), and recent results suggest that the two-step induction mechanism found in O. tipulae is ancestral (Kiontke et al., in preparation).
|
Figure 3. Vulva development in Oscheius tipulae. Left panel. Successive stages of vulval development. Differences with C. elegans are highlighted in green. AC: anchor cell. S: syncytial fate. U: undivided granddaughter, characteristic of 2° vulval fate. T: transverse (left-right division), characteristic of 1° vulval fate. Right panel. A-C. Nomarski pictures of corresponding stages. In (C), the four granddaughters of P5.p and P7.p are easily visible in the same focal plane on either side of the vulval invagination (stars next to their nuclei). The eight progeny of P6.p are out of focus. D. MH27 staining of cell junctions at the late L4 stage; the uterus cell junctions are visible dorsally to the vulva (top) (courtesy of I. Kolotuev).
O. tipulae has been used for extensive genetic screens for Egl (Egg-laying defective) and Pvl (Protruding vulva) mutants (Dichtel-Danjoy and Félix, 2004; Sommer, 2000). Those with abnormal vulva development were divided in three categories corresponding to successive steps in vulval development:
Vulva competence group formation and centering : Cov mutants (for Competence/centering Of the Vulva), including a deletion in Oti-lin-39 (Louvet-Vallée et al., 2003);
Vulval fate induction: Iov (for Induction Of the Vulva; Dichtel-Danjoy and Félix, 2004);
Division of vulval precursor cells, without affecting their vulval fates: Dov (for Division Of the Vulva; Dichtel et al., 2001).
Interestingly, the spectrum of phenotypes obtained in vulva mutant screens in O. tipulae strongly differs from that obtained in C. elegans. For example, the Dov mutations have no equivalent in C. elegans, or are at least not found at the same rate. In contrast, Hypoinduced phenotypes have proven difficult to isolate in O. tipulae. Also, the 1°/2° fate pattern of Hyperinduced mutants differ between the two species. This comparison of parallel mutagenesis screens on a homologous developmental system shows that the spectrum of phenotypes that can be reached by random mutagenesis evolves (Dichtel-Danjoy and Félix, 2004).
A particularly interesting phenotype of Oscheius species is the number of division rounds of P4.p and P8.p: zero in Oscheius guentheri, one in O. dolichura and O. dolichuroides (as in C. elegans), but usually two in O. tipulae, spp. 2 and 3. However, whereas P4.p and P8.p divide twice in the CEW1 reference strain of O. tipulae, this trait is highly polymorphic within an isogenic strain or between wild isolates of O. tipulae (Delattre and Félix, 2001a; Delattre and Félix, 2001b). From the analysis of recombinant inbred lines between the two isolates CEW1 and PS959, the difference between them (two versus one division round) is caused by variations at several segregating loci (Delattre and Félix, 2001a). P(4,8).p division is a trait that is easily mimicked by mutation in CEW1, as shown by the isolation of several mutants with one or zero division round of these cells (Dichtel et al., 2001).
I thank M.-L. Dichtel-Danjoy, M. Delattre, S. Louvet-Vallée, G. Maro and D. Baïlle for published and unpublished work on Oscheius and K. Kiontke and anonymous reviewers for comments.
Ahn, I.-Y., and Winter, C.E. (2005). Determination of DNA base composition by small scale acrylamide-CsCl gradient centrifugation. J. Biochem. Biophys. Methods 63, 155–160. Abstract Article
Barrière, A., and Félix, M.-A. (2005). High local genetic diversity and low outcrossing rate in Caenorhabditis elegans natural populations. Curr. Biol. 15, 1176–1184. Abstract Article
Blaxter, M.L., De Ley, P., Garey, J.R., Liu, L.X., Scheldeman, P., Vierstraete, A., Vanfleteren, J.R., Mackey, L.Y., Dorris, M., Frisse, L.M., et al. (1998). A molecular evolutionary framework for the phylum Nematoda. Nature 392, 71–75. Abstract Article
Brenner, S. (1974). The genetics of Caenorhabditis elegans. Genetics 77, 71–94. Abstract
Delattre, M., and Félix, M.-A. (2001a). Microevolutionary studies in nematodes: a beginning. Bioessays 23, 807–819. Abstract Article
Delattre, M., and Félix, M.-A. (2001b). Polymorphism and evolution of vulval precursor cell lineages within two nematode genera, Caenorhabditis and Oscheius. Curr. Biol. 11, 631–643. Abstract Article
Delattre, M., and Félix, M.-A. (2001c). Evolution and development of a variable left-right asymmetry in nematodes: the handedness of P11/P12 migration. Dev. Biol. 232, 362–371. Abstract Article
Dichtel, M.-L., Louvet-Vallée, S., Viney, M.E., Félix, M.-A., and Sternberg, P.W. (2001). Control of vulval cell division number in the nematode Oscheius/Dolichorhabditis sp. CEW1. Genetics 157, 183–197. Abstract
Dichtel-Danjoy, M.-L., and Félix, M.-A. (2004). Phenotypic neighborhood and micro evolvability. Trends Genet. 20, 268–276. Abstract Article
Dichtel-Danjoy, M.-L., and Félix, M.-A. (2004). The two steps of vulval induction in Oscheius tipulae CEW1 recruit common regulators including a MEK kinase. Dev. Biol. 265, 113–126. Abstract Article
Evans, D., Zorio, D., MacMorris, M., Winter, C.E., Lea, K., and Blumenthal, T. (1997). Operons and SL2 trans-splicing exist in nematodes outside the genus Caenorhabditis. Proc. Natl. Acad. Sci. U.S.A. 94, 9751–9756. Abstract Article
Félix, M.-A., Delattre, M., and Dichtel, M.-L. (2000). Comparative developmental studies using Oscheius/Dolichorhabditis sp. CEW1 (Rhabditidae). Nematology 2, 89–98. Article
Félix, M.-A., and Sternberg, P.W. (1996). Symmetry breakage in the development of one-armed gonads in nematodes. Development 122, 2129–2142. Abstract
Félix, M.-A., and Sternberg, P.W. (1997). Two nested gonadal inductions of the vulva in nematodes. Development 124, 253–259. Abstract
Félix, M.-A., Vierstraete, A., and Vanfleteren, J. (2001). Three biological species related to Rhabditis (Oscheius) pseudodolichura Körner in Osche, 1952. J. Nematol. 33, 104–109.
Flemming, A.J., Shen, Z.-Z., Cunha, A., Emmons, S.W., and Leroi, A.M. (2000). Somatic polyploidization and cellular proliferation drive body size evolution in nematodes. Proc. Natl. Acad. Sci. U.S.A. 97, 5285–5290. Abstract Article
Lam, A.B.Q., and Webster, J.M. (1971). Morphology and biology of Panagrolaimus tipulae n. sp. (Panagrolaimidae) and Rhabditis (Rhabditella) tipulae n. sp. (Rhabditidae), from leatherjacket larvae, Tipula paludosa (Diptera: Tipulidae). Nematologica 17, 201–212.
Li, X., Massey, H.C., Jr., Nolan, T.J., Schad, G.A., Kraus, K., Sundaram, M., and Lok, J.B. (2006). Successful transgenesis of the parasitic nematode Strongyloides stercoralis requires endogenous non-coding control elements. Int. J. Parasitol. Abstract Article
Louvet-Vallée, S., Kolotuev, I., Podbilewicz, B., and Félix, M.-A. (2003). Control of vulval competence and centering in the nematode Oscheius sp. 1 CEW1. Genetics 163, 133–146. Abstract
Sommer, R.J. (2000). Comparative genetics: A third model nematode species. Curr. Biol. 10, R879–R881. Abstract Article
Sommer, R.J., and Sternberg, P.W. (1995). Evolution of cell lineage and pattern formation in the vulval equivalence group of rhabditid nematodes. Dev. Biol. 167, 61–74. Abstract Article
Sudhaus, W. (1976). Vergleichende Untersuchungen zur Phylogenie, Systematik, Ökologie, Biologie und Ethologie der Rhabditidae (Nematoda). Zoologica 43, 1–228.
Sudhaus, W. (1993). Redescription of Rhabditis (Oscheius) tipulae (Nematoda: Rhabditidae) associated with leatherjackets, larvae of Tipula paludosa (Diptera: Tipulidae). Nematologica 39, 234–239.
Sudhaus, W., and Hooper, D.J. (1994). Rhabditis (Oscheius) guentheri sp.n., an unusual species with reduced posterior ovary, with observations on the Dolichura and Insectivora groups (Nematoda: Rhabditidae). Nematologica 40, 508–533.
Winter, C.E. (1992). The yolk polypeptides of a free-living rhabditid nematode. Comp. Biochem. Physiol. 103B, 189–196.
Winter, C.E., Penha, C., and Blumenthal, T. (1996). Comparison of a vitellogenin gene between two distantly related rhabditid nematode species. Mol. Biol. Evol. 13, 674–684. Abstract
*Edited by Jonathan Hodgkin. Last revised June 30, 2006. Published August 16, 2006. This chapter should be cited as: Félix, M.-A. Oscheius tipulae (August 16, 2006), WormBook, ed. The C. elegans Research Community, WormBook, doi/10.1895/wormbook.1.119.1, http://www.wormbook.org.
Copyright: © 2006 Marie-Anne Félix. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
§To whom correspondence should be addressed.
 All WormBook content, except where otherwise noted, is licensed under a Creative Commons Attribution License.
All WormBook content, except where otherwise noted, is licensed under a Creative Commons Attribution License.