Embryo series courtesy of Einhard Schierenberg
Embryo series courtesy of Einhard SchierenbergTable of Contents
Abstract
The C. elegans embryo is a powerful model system for studying the mechanics of metazoan cell division. Its primary advantage is that the architecture of the syncytial gonad makes it possible to use RNAi to generate oocytes whose cytoplasm is reproducibly (typically >95%) depleted of targeted essential gene products via a process that does not depend exclusively on intrinsic protein turnover. The depleted oocytes can then be analyzed as they attempt their first mitotic division following fertilization. Here we outline the characteristics that contribute to the usefulness of the C. elegans embryo for cell division studies. We provide a timeline for the first embryonic mitosis and highlight some of its key features. We also summarize some of the recent discoveries made using this system, particularly in the areas of nuclear envelope assembly/ dissassembly, centrosome dynamics, formation of the mitotic spindle, kinetochore assembly, chromosome segregation, and cytokinesis.
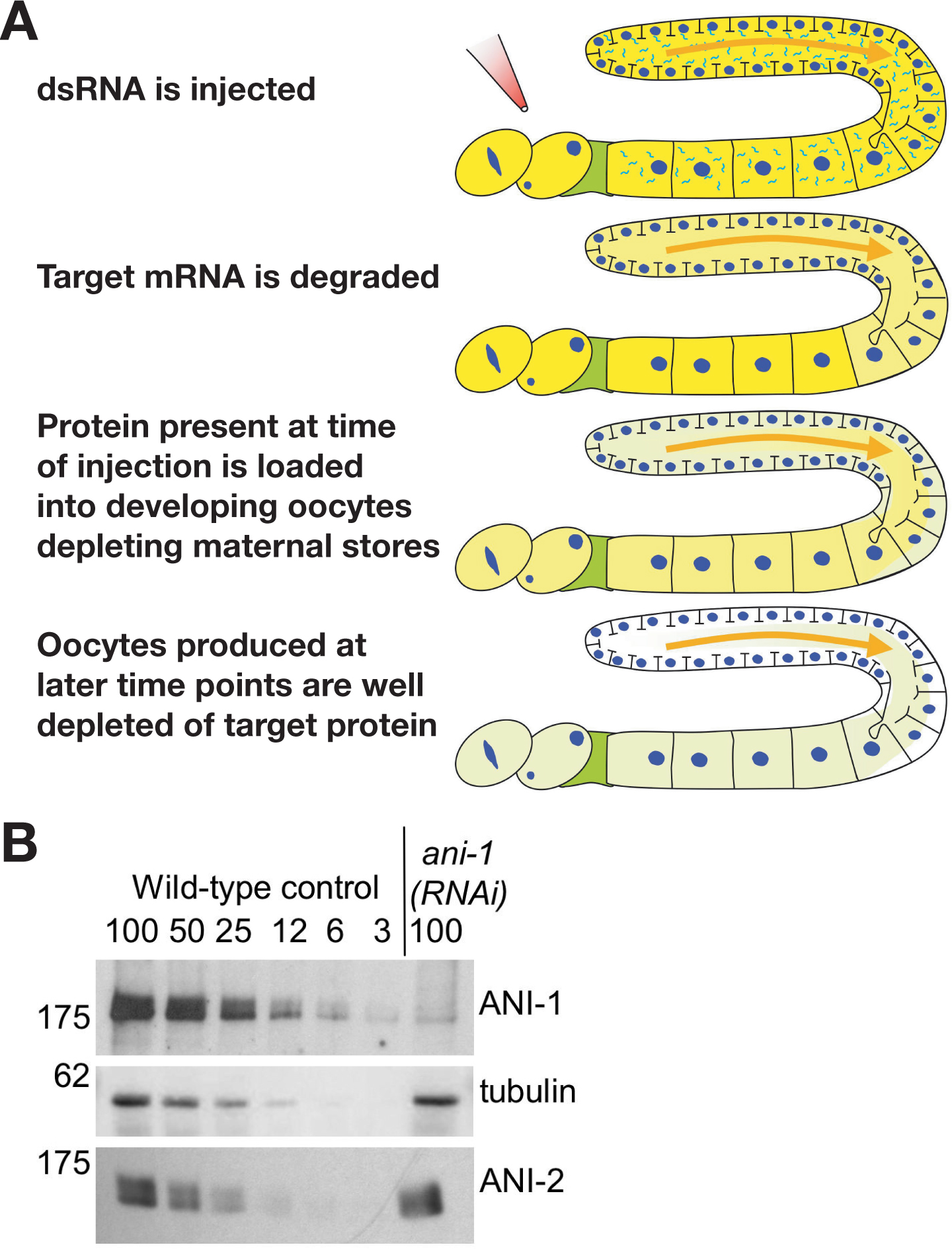
The C. elegans embryo is a powerful model system for studying the mechanics of metazoan cell division. Its primary advantage is that the syncytial gonad makes it possible to use RNA interference (RNAi) to generate oocytes whose cytoplasm is reproducibly (>95%) depleted of targeted essential gene products. Introduction of dsRNA rapidly catalyzes the destruction of the corresponding mRNA in many different systems. However, depletion of pre-existing protein is generally a slow process that depends on the half-life of the targeted protein. In contrast, in the C. elegans gonad, the protein present when the dsRNA is introduced is depleted by the continual packaging of maternal cytoplasm into oocytes (Figure 1). Since depletion relies on the rate of embryo production instead of protein half-life, the kinetics tend to be similar for different targets. By 36-48 hours after introduction of the dsRNA, newly formed oocytes are typically >95% depleted of the target protein.
|
Figure 1. The generation of oocytes depleted of target proteins by RNAi in C. elegans does not require intrinsic protein turnover. (A) The gonad is a syncytial tube lined with nuclei in various stages of meiotic prophase. The meiotic nuclei contribute mRNA that is translated to generate the protein that is loaded into the developing oocytes. The meiotic nuclei and all of the developing oocytes except the final 4-5 (Maddox et al., 2005) are diffusionally connected to the rachis (the central cytoplasmic core of the syncytial gonad). Introduction of a dsRNA triggers the degradation of the corresponding mRNA. However, the syncytial gonad and connected oocytes still contain the target protein that was present at the time of injection. Maternal stores are depleted by the continual packaging of gonad cytoplasm into developing oocytes. By 36 to 48 hours after introduction of the dsRNA oocytes are typically >95% depleted of the target protein. (B) Quantitative western blot of worms injected with a dsRNA against ANI-1, a protein required for cortical contractility in the early embryo (Maddox et al., 2005). Serial dilutions of identically processed control worms were loaded to quantify the depletion level. ANI-1 is ~97% depleted in the injected worms, whereas levels of two control proteins, α-tubulin and a related actin binding protein, ANI-2, are unaffected. Figure courtesy of Amy Maddox.
Several additional advantages contribute to the usefulness of the C. elegans embryo as a model system. Of particular importance is their rapid and highly stereotypical mitotic divisions; the time between the onset of DNA condensation and the completion of furrow ingression during cytokinesis is approximately 14 minutes. The invariant nature of the first few divisions also facilitates the development of methods to assess the consequences of molecular perturbations. Quantitative methods have already been developed to monitor pronuclear migration (Figure 4; see also Albertson, 1984; O'Connell, 2000), cortical flows (for examples see Cheeks et al., 2004; Hird and White, 1993; Munro et al., 2004), chromosome segregation and spindle elongation during anaphase (for examples see Cheeseman et al., 2004; Grill et al., 2001; Labbe et al., 2004), and the asymmetric positioning of the spindle within the embryo (for examples see Colombo et al., 2003; Labbe et al., 2004; Tsou et al., 2002). Assay development has been accelerated by the emergence of microparticle bombardment mediated transformation (Praitis et al., 2001) and the availability of vectors containing regulatory sequences that direct efficient germline expression (Strome et al., 2001), which together have led to the generation of a large number of strains expressing fluorescent fusion proteins in the early embryo. Analysis of the mechanical consequences of depleting essential cell division proteins is also facilitated by the relatively weak DNA damage (Brauchle et al., 2003) and spindle checkpoints (Encalada et al., 2004), which allow the embryo to proceed through the cell cycle despite dramatic defects in nuclear structure, spindle assembly, chromosome segregation and centrosome function.
Genetic and RNAi-based approaches have identified a large number of loci important for cell division. Mutants in proteins required for cell division have been uncovered in screens of collections of nonconditional maternal effect and temperature sensitive mutations that result in embryonic lethality (for some examples see Encalada et al., 2000; Golden et al., 2000; Gönczy et al., 1999; Kemphues et al., 1988; O'Connell et al., 1998). The ability to reproducibly deplete oocytes of target proteins by RNAi, which can be performed by feeding, soaking or injection of hermaphrodites (see Reverse genetics), has also led to a series of genome-wide screens that identified a set of ~2100 genes required for embryonic viability (Fernandez et al., 2005; Fraser et al., 2000; Gönczy et al., 2000; Kamath et al., 2003; Maeda et al., 2001; Piano et al., 2000; Rual et al., 2004; Simmer et al., 2003; Sõnnichsen et al., 2005). Filming of embryos depleted of each of these 2100 gene products using differential interference contrast (DIC) microscopy has defined a set of 660 genes whose inhibition results in detectable defects during the first two cell divisions (Gönczy et al., 2000; Piano et al., 2000; Sõnnichsen et al., 2005; Zipperlen et al., 2001). Roughly half of these genes are specifically required for cell division processes such as chromosome segregation or cytokinesis, whereas the other half contributes to cell maintenance, via roles in processes such as translation and mitochondrial function (Sõnnichsen et al., 2005). The embryonic lethality resulting from RNAi of some of the 1440 genes for which no DIC defect was observed may be due to cell division defects that remain undetected, either due to incomplete penetrance of the RNAi or failure to score the resulting defects by DIC (for example subtle defects in chromosome segregation are very difficult to detect using this assay). Alternatively, depletion of many of these gene products may cause embryonic lethality due to developmental defects that preclude hatching.
Due to its accessibility to RNAi-based molecular perturbation, the first embryonic division that ensues following fertilization has been the most intensively studied. In this section, we provide a brief timeline for the events between fertilization and the completion of the first cytokinesis (outlined schematically in Figure 2; for reviews see Cowan and Hyman, 2004; Pelletier et al., 2004; Schneider and Bowerman, 2003).
Prior to fertilization, C. elegans oocytes are arrested in meiotic prophase with nuclei containing two copies of the diploid genome packaged into recombined bivalent chromosomes. The two rounds of meiotic chromosome segregation that generate the haploid oocyte pronucleus are completed in the zygote after the oocytes are fertilized. During each meiotic division, chromosome segregation is accomplished by a small acentriolar meiotic spindle that forms in the embryo anterior. During anaphase of meiosis I and again in meiosis II, the meiotic spindle associates with the cortex in an end-on fashion, and a highly asymmetric cytokinesis-like event extrudes a polar body (Figure 2; Albertson and Thomson, 1993; Clark-Maguire and Mains, 1994; Yang et al., 2003). In addition to the haploid pronucleus, the sperm brings a pair of centrioles into the oocyte, which lacks centrioles due to their degradation during oogenesis. As meiosis completes, the haploid oocyte and sperm-derived pronuclei, located at opposite ends of the embryo increase in size, becoming visible by DIC microscopy. After entering the oocyte, the sperm-derived centriole pair recruits pericentriolar material and acquires the ability to nucleate microtubules (O'Connell, 2000; Pelletier et al., 2004). Subsequently, the two sperm-derived centrioles separate, forming two centrosomes positioned on either side of the paternal pronucleus. Coincident with chromosome condensation during mitotic prophase, the pronuclei migrate towards each other. After the pronuclei meet, the nuclear-centrosome complex moves to the center of the embryo and rotates to align with the long axis of the embryo (Albertson, 1984; Hyman and White, 1987). The miotitc spindle begins to move towards the embryo posterior during metaphase (Labbe et al., 2004; Oegema et al., 2001), and asymmetric elongation during anaphase contributes to its posterior displacement (Albertson, 1984; Grill et al., 2001). Since the cleavage furrow bisects the mitotic spindle, this displacement results in an asymmetric first cleavage (For more on the mechanisms that generate this asymmetry see Asymmetric cell division and axis formation in the embryo).
|
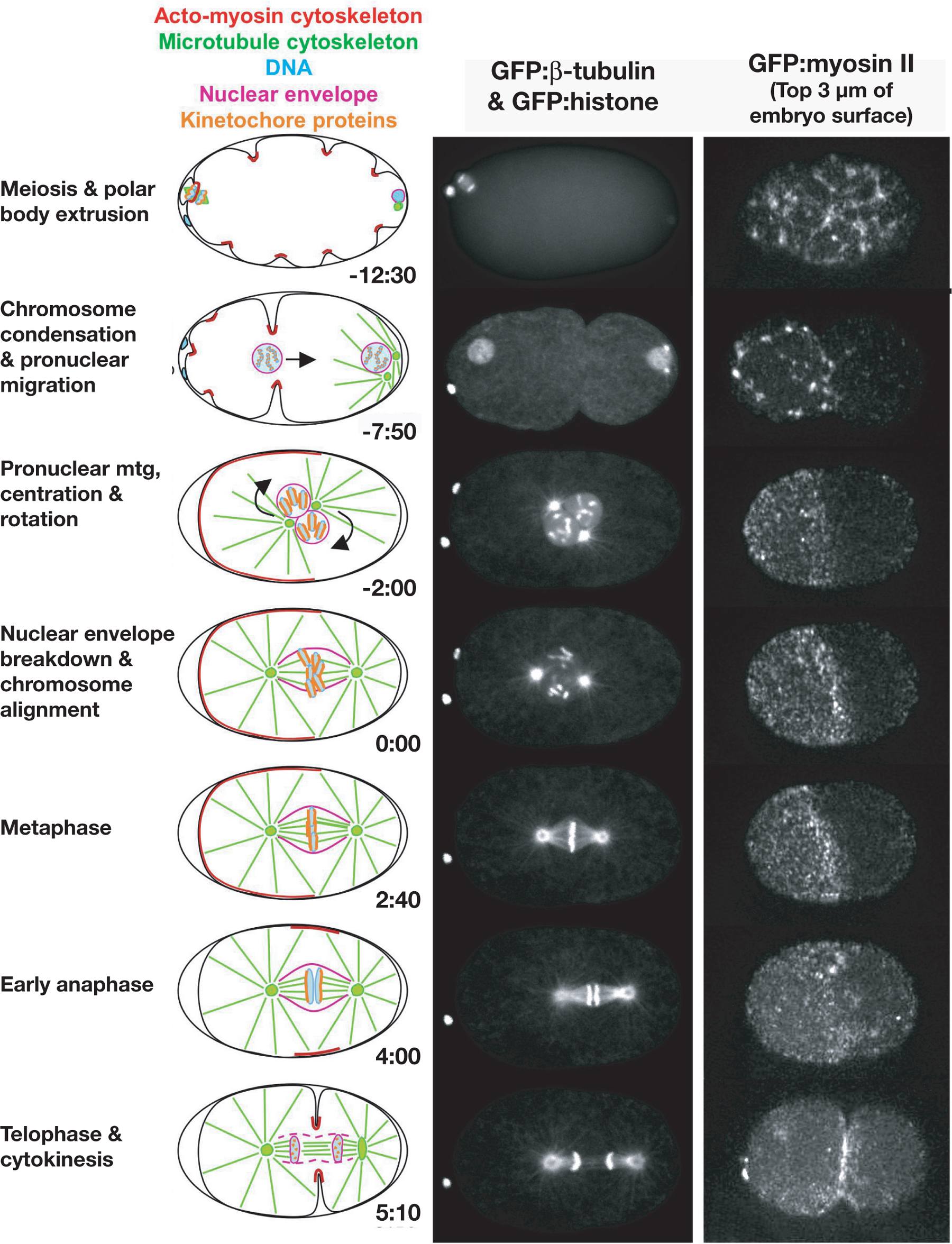
Figure 2. Nuclear envelope dynamics in the C. elegans embryo. (Left column) Schematics illustrate the major features of the first division. Approximate times are in minutes:seconds after nuclear envelope breakdown. (Middle column) Images of each stage in a strain expressing GFP: beta-tubulin and GFP: histone to simultaneously visualize the microtubule cytoskeleton and the DNA. The top image of anaphase of meiosis II was taken from a movie collected by wide-field microscopy. All subsequent images are of the same embryo and were collected by spinning disk confocal microscopy (images courtesy of Carrie Cowan). (Right column) Stills from a timelapse sequence of an embryo expressing GFP:myosin II (images courtesy of Amy Maddox; strain provided by Ed Munro). For each time point, three spinning disk confocal images of the embryo surface were collected at 1 μm intervals and projected. During polar body extrusion, ruffles form over the entire cortex. Foci of myosin II are apparent at the base of each of the ingressing ruffles. As polarity is established, myosin II concentrates in an anterior cortical cap that persists into metaphase (Munro et al., 2004). During cytokinesis, an equatorial band of cortical myosin II forms in the plane defined by the spindle midzone. Figure courtesy of Amy Maddox.
The C. elegans nuclear envelope is structurally similar to that of vertebrates, consisting of two concentric membranes (outer and inner) enclosing a lumenal space, nuclear pore complexes that mediate bidirectional transport between the cytoplasm and the nucleus, and an underlying laminar network (Figure 3; Cohen et al., 2002). The molecular composition of the C. elegans inner nuclear membrane/lamina also resembles that in vertebrates (Table 1). C. elegans expresses a single B-type lamin (LMN-1; Liu et al., 2000; Riemer et al., 1993), that forms a meshwork of intermediate filaments beneath the inner nuclear membrane (reviewed in Gruenbaum et al., 2005). The C. elegans inner nuclear membrane/lamina also contains three proteins, Ce-emerin, CeMAN-1, and CeLem2, that contain a LEM domain, a defining 40 amino acid motif shared by a family of nuclear envelope proteins (Gruenbaum et al., 2002; Lee et al., 2000; Lin et al., 2000; Liu et al., 2003). LEM family proteins all bind to lamins (reviewed in Lee and Wilson, 2004) and to the small inner nuclear membrane associated protein BAF (Segura-Totten and Wilson, 2004; Zheng et al., 2000). Depletion of LMN-1, Ce-BAF, or simultaneous depletion of the LEM family proteins Ce-Emerin and Ce-MAN-1, results in a similar spectrum of defects in nuclear structure, chromosome condensation, and chromosome segregation (Liu et al., 2000; Liu et al., 2003; Margalit et al., 2005; Zheng et al., 2000).
Table 1. Nuclear envelope proteins
| C. elegans protein | C. elegans gene | Required for embryonic viability? | Vertebrate orthologue | TMR | Summary of localization and functional analysis | Selected references |
|---|---|---|---|---|---|---|
| Ce-lamin (LMN-1) | lmn-1 (DY3.2) | YES | Lamin-B | NO | B-type lamin expressed in all cell types; localizes to the nuclear side of the nuclear envelope; required for nuclear morphology, nuclear pore complex distribution, chromosome segregation and the localization of Ce-emerin. | Liu, 2000; Gruenbaum, 2002 |
| Ce-emerin (EMR-1) | emr-1 (M01D7.6) | NO | Emerin | YES | Integral component of the inner nuclear membrane; contains a LEM domain and interacts with Ce-Lamin; co-depletion with Ce-MAN-1 results in 100% embryonic lethality, chromosome condensation and segregation defects and mislocalization of Ce-BAF. | Gruenbaum, 2002; Liu, 2003 |
| Ce-MAN1 (LEM-2) | lem-2 (W01G7.5) | PARTIAL Depletion gives 15% lethality | LEM2 and MAN1 | YES | Integral component of the inner nuclear membrane; contains a LEM domain and interacts with Ce-Lamin and Ce-BAF in vitro. May require Ce-Lamin for its localization. | Liu, 2003 |
| Ce-BAF | baf-1 (B0464.7) | YES | BAF | NO | 10kDa soluble protein highly conserved among metazoans; localization to the NE requires emerin and MAN1; interacts with Ce-MAN1 in vitro; required for the proper organization of Ce-lamin, Ce-emerin and Ce-MAN1 on the nuclear envelope. | Liu, 2003; Margalit, 2005 |
| Matefin SUN-1 | mtf-1 sun-1 (F57 B1.2) | YES | None | NO | Interacts with lamin, but its Localization to the nuclear envelope does not require lamin; expressed in all embryonic cells until mid-embryogenesis and thereafter only in germline cells; required for ZYG-12 localization and the attachment of centrosomes to nuclei. | Fridkin, 2004; Malone, 2003 |
| ZYG-12 | zyg-12 (ZK546.1) | YES | SYNE | YES/NO* | Member of the Hook family of proteins; localizes to the nuclear envelope and accumulates around the centrosomes; required for the attachment of centrosomes to nuclei; * TMR present in some isoforms. | Malone, 2003 |
| UNC-83 | unc-83 (W01A11.3) | NO | ? | NO | Requires UNC-84 for its localization to the nuclear envelope; mutations disrupt nuclear migration in migrating P cells, hyp7 precursors, and the intestinal primordium. | Starr, 2001 |
| UNC-84 | unc-84 (F54B11.3) | NO | UNC84 | YES | Integral nuclear envelope protein required for nuclear migration and anchorage; expressed after the 26-cell stage; localization requires lamin; required to target UNC-83 and ANC-1, which function in nuclear migration and positioning, respectively, to the nuclear membrane; during nuclear positioning, may form a bridging complex with ANC-1 that spans the perinuclear space to connect the NE to actin cytoskeleton. | Starr, 2001; Malone, 1999; Lee, 2002; Starr, 2002 |
| ANC-1 | anc-1 (ZK973.6) | NO | Nesprin/ Nuance | YES | 955kDa protein orthologous to vertebrate NUANCE proteins; connects the nuclear envelope to the actin cytoskeleton during nuclear positioning in a UNC-84-dependent manner. | Starr, 2002 |
The dynamics of nuclear envelope disassembly and reassembly during the first mitotic division of the C. elegans embryo are illustrated in Figure 3. LMN-1 leaves the nuclear envelope during prometaphase (Lee et al., 2000; Liu et al., 2000). In contrast, inner nuclear membranes containing Ce-emerin and Ce-MAN-1 remain largely intact and surround the mitotic spindle everywhere except near spindle poles during metaphase and early anaphase, disassembling fully only during mid to late anaphase (Figure 3; Lee et al., 2000; Lee et al., 2002; V. Galy, P. Askjaer and I.W. Mattaj, personal communication). As the remnants of the old nuclear envelopes disperse, the formation of new nuclear envelopes around the segregated chromatin is detected beginning about 1 minute after anaphase onset (Figure 3; V. Galy, P. Askjaer and I.W. Mattaj, personal communication).
|
Figure 3. Nuclear envelope dynamics in the C. elegans embryo. Figure courtesy of Vincent Galy. (A) Schematics illustrate the cycle of nuclear envelope breakdown and reassembly. (B) Still images of the first mitotic division of wild-type embryos expressing YFP-Lamin (left) and GFP-MAN1 (right). Times on the right are relative to first metaphase to anaphase transition. Arrowheads indicate the reappearance of GFP-MAN1 around the chromatin at t=60 sec and YFP-Lamin at t=120 sec. White stars mark the positions of the centrosomes. Note the persistence of membranes containing GFP-MAN1 around the mitotic spindle and centrosomes. Scale bar = 10μm.
The molecular composition of the nuclear pore complexes (NPCs) is also similar to that in vertebrates. C. elegans orthologs of at least one component of each vertebrate NPC sub-complex have been identified (Galy et al., 2003; Kuznetsov et al., 2002). 17 genes encoding 19 nucleoporins are essential for embryonic viability. Depletion of 14 of these proteins results in defects in nuclear morphology and, in some cases, to reduced nuclear size consistent with a defect in nucleo-cytoplasmic transport (Galy et al., 2003). Nuclear envelope assembly also requires the small GTPase, Ran and the nuclear transport receptor importin-γ (IMB-1; Askjaer et al., 2002; Bamba et al., 2002; Walther et al., 2003). High concentrations of RanGTP in the vicinity of chromatin are thought to promote the dissociation of importin-γ from nucleoporins to trigger NPC assembly on the chromatin surface.
The site of sperm entry defines the embryo posterior (Goldstein and Hird, 1996). As the pronuclei become visible by DIC, the sperm-derived pronucleus and its associated centrosome(s) sit on the posterior cortex. The oocyte-derived pronucleus forms following two rounds of meiotic chromosome segregation, typically in the embryo anterior. The two pronuclei migrate towards each other coincident with chromosome condensation during the first mitotic prophase. Pronuclear migration consists of movement of the oocyte pronucleus towards the sperm pronucleus and movement of the sperm pronucleus away from the cortex towards the embryo center (Albertson, 1984; O'Connell et al., 2000). Initially, the oocyte pronucleus moves ~ 12 μm towards the posterior at a slow rate (~ 3.5 μm/min). As it approaches the sperm pronucleus, the oocyte pronucleus accelerates, moving an additional 10 μm at ~5-10 times its initial rate (Figure 4; Albertson, 1984; O'Connell et al., 2000). The sperm pronucleus begins its migration later than its female counterpart and travels at a slow rate of ~ 3.5 μm/min till it meets the oocyte pronucleus near the embryo center (~7 μm).
|
Figure 4. Kinetics of pronuclear migration in the C. elegans embryo. Timelapse DIC sequences of 20 wild-type and 16 gamma-tubulin depleted embryos were collected. The average position of the oocyte-derived and sperm-derived pronuclei along the anterior-posterior axis of the embryo is plotted (x-axis) as a function of time (y-axis). Times are with respect to pronuclear meeting. The sperm pronucleus moves towards the embryo anterior at a uniform slow rate. The oocyte pronucleus initially moves towards the embryo posterior at a similar slow rate (Slow phase), but then speeds up prior to nuclear meeting (Fast phase). Pronuclear migration depends on the timing of formation and size of centrosomal microtubule asters. In embryos, depleted of gamma-tubulin centrosomal microtubule asters form later than in wild-type (Hannak et al., 2002) and the fast phase of pronuclear migration is not observed. A similar phenotype has been characterized in embryos mutant for the centrosomal protein SPD-2, in which the centrosomal microtubule asters are highly attenuated (O'Connell et al., 2000). Figure courtesy of Eva Hannak and Stephan Grill.
Rapid movement of the oocyte pronucleus towards the sperm pronucleus and pronuclear meeting, both require an intact microtubule cytoskeleton (Strome and Wood, 1983). Two nuclear envelope proteins, ZYG-12 and SUN-1 recruit dynein to pronuclei, and are required for centrosomes to maintain their association with nuclei (Table 1; Malone et al., 2003). The centrosomes separate around the sperm pronucleus in a dynein dependent manner (Gönczy et al., 1999). As the pronuclei move towards each other, dynein on the oocyte pronucleus is thought to come into contact with microtubules emanating from the centrosomal asters associated with the sperm pronucleus. The fast phase of pronuclear migration and pronuclear meeting is thus mediated by nuclear envelope associated dynein pulling on the two centrosomal microtubule asters (Cowan and Hyman, 2004; Gönczy et al., 1999; Hamill et al., 2002; Malone et al., 2003; O'Connell et al., 2000; Schmidt et al., 2005).
In addition to the migration of the female pronucleus towards the male pronucleus, the migration and centration of the male pronucleus within the embryo has also been analyzed to distinguish between two possible models: (1) a “pushing mechanism,” in which the male pronucleus is pushed away from the cortex by the polymerization of astral microtubules and, (2) a “pulling mechanism” in which the male pronucleus is pulled by minus-end-directed motors anchored throughout the cytoplasm. Comparisons between simulations and actual migration indicate that the second “pulling” model is the primary mechanism (Kimura and Onami, 2005). Although microtubule-based mechanisms for pronuclear movement are the best characterized, some slow pronuclear movement is still observed in embryos in which the microtubule cytoskeleton is compromised, or which lack centrosomes or fail to recruit dynein to nuclei (Cowan and Hyman, 2004; Gönczy et al., 1999; Hamill et al., 2002; Malone et al., 2003; O'Connell et al., 2000; Schmidt et al., 2005). Consistent with these observations, correlative evidence suggests that cortical flows may also contribute to the slow phase of pronuclear migration (Hird and White, 1993).
Centrosomes consist of a single centriole or centriole pair surrounded by pericentriolar material that nucleates and anchors microtubules. Like the centrioles in Drosophila embryos (Callaini and Riparbelli, 1990), C. elegans centrioles are composed of nine singlet microtubules symmetrically positioned around a central tube (Figure 5B; Albertson, 1984; Kirkham et al., 2003; O'Connell et al., 2001; Wolf et al., 1978). Each cylindrical centriole is approximately 200– 250 nm in length and 175 nm in diameter. In contrast, vertebrate centrioles typically have nine triplet microtubules (Marshall, 2001; Preble et al., 2000). Consistent with this structural difference, the C. elegans genome lacks homologs of delta- and epsilon-tubulin, two tubulin family members required for the formation of triplet microtubules (Dutcher, 2003).
|
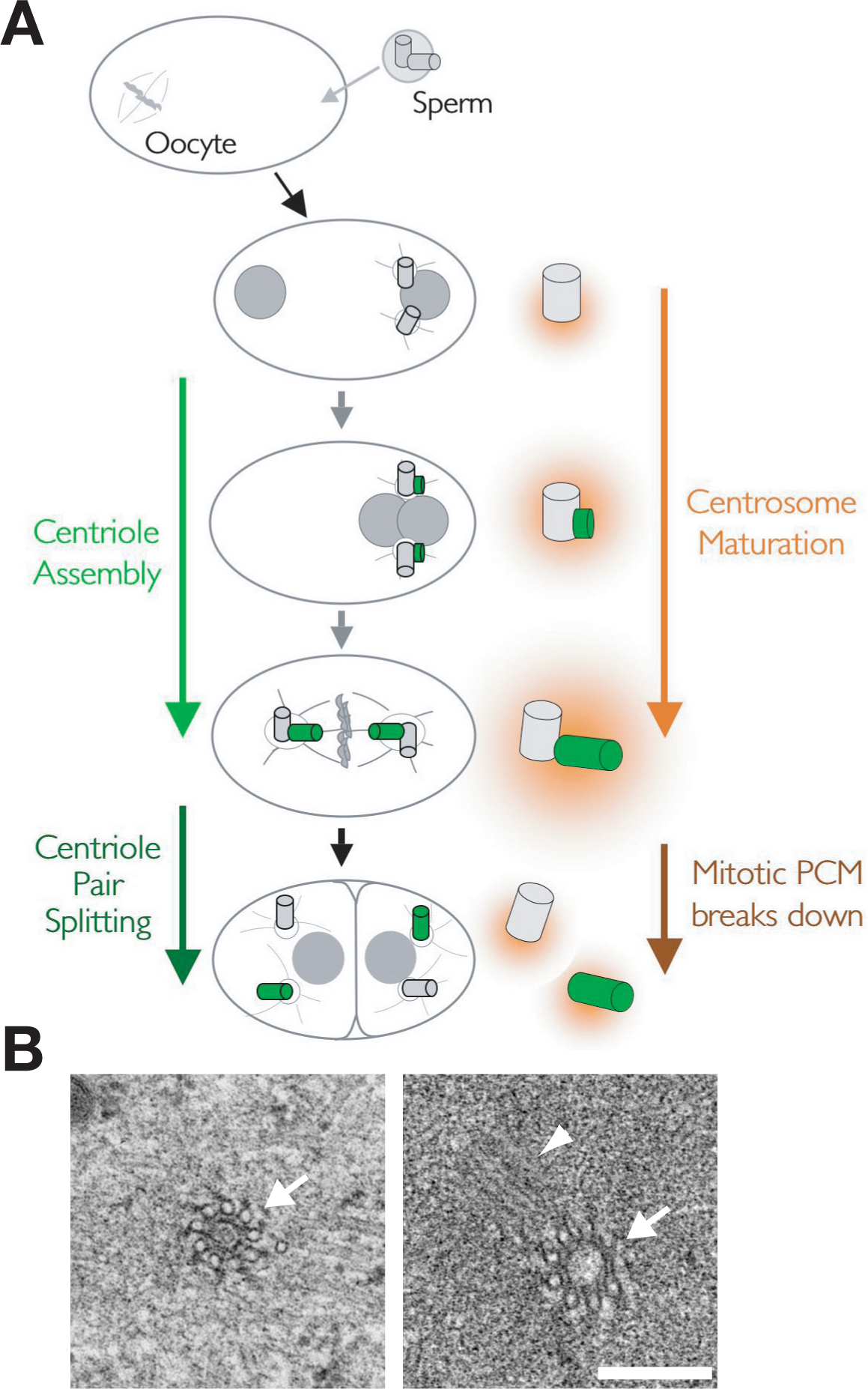
Figure 5. Mitotic kinetochores in the C. elegans embryo. (A) Schematic of the first cycle of centrosome duplication that immediately follows fertilization. Figure and electron micrographs courtesy of Amy Maddox and Thomas Müller-Reichert, respectively. Centrosome duplication consists of alternating cycles of new centriole assembly and splitting of the centriole pairs. A pair of centrioles (grey) enters the egg with the sperm during fertilization. The sperm centrioles acquire pericentriolar material (PCM; orange) in the egg and begin to nucleate microtubule asters. New daughter centrioles (green) assemble adjacent to each of the sperm centrioles so that by metaphase each centrosome contains two full-length centrioles, one inherited from the sperm and one that formed in the embryo cytoplasm. As the embryo enters mitosis, the amount of PCM around the centrioles and the number of microtubules nucleated by the centrosomes increases in a process called centrosome maturation. The centriole pairs split in late anaphase/telophase and the mitotic PCM breaks down, so that each daughter cell inherits a pair of small centrosomes each containing a single centriole. (B) Two images of prometaphase/metaphase centrioles with singlet microtubules in either cross section (arrows) or a longitudinal orientation (arrowhead) are shown. Bar is 250 nm.
During fertilization, the amoeboid sperm brings a pair of centrioles into the egg. After entering the egg, the centrioles recruit pericentriolar material and separate around the sperm-derived nucleus, forming two centrosomal microtubule asters (Figure 5; O'Connell, 2000; Pelletier et al., 2004). After they separate, a new daughter centriole begins to form adjacent to each sperm centriole. The centrosomes recruit additional pericentriolar material as the embryo enters mitosis in a process called centrosome maturation, increasing about 5 fold in size and nucleating capacity by metaphase (Hannak et al., 2001). Coincident with the recruitment of additional PCM, the daughter centrioles elongate, reaching full length by metaphase. In late anaphase/telophase the centriole pairs split. Concurrent with cytokinesis, the mitotic PCM disassembles, releasing two small centrosomes into each daughter cell.
Pericentriolar material is thought to consist of a proteinacious matrix, called the “centromatrix”, that recruits other PCM components (Bornens, 2002; Palazzo et al., 2000). Three centrosomal proteins, SPD-5, SPD-2 and AIR-1 (Aurora-A kinase), are required for the assembly of pericentriolar material (see Table 2 for a list of C. elegans centrosome/centriole components). SPD-5 is a coiled-coil protein thought to be a component of the centromatrix. In spd-5 embryos, no PCM forms around the centrioles, and no centrosomal microtubule asters are observed (Hamill et al., 2002). Severe defects in PCM assembly are also observed when SPD-2 or the aurora A kinase homolog AIR-1 are inhibited (Hannak et al., 2001; Kemp et al., 2004; O'Connell et al., 2000; Pelletier et al., 2004; Schumacher et al., 1998). In these embryos, centrosomal microtubule asters form, but are very small, and additional PCM fails to accumulate around the centrioles during mitotic entry.
Table 2. Centrosome proteins
| C. elegans protein | C. elegans gene | Required for embryonic viability? | Vertebrate orthologue | Summary of localization and functional analysis | Selected references |
|---|---|---|---|---|---|
| SAS-4 | sas-4 (F10E9.8) | YES | CPAP | Centriole protein required for centriole assembly; incorporated into centrioles during their assembly and does not subsequently exchange with the cytoplasmic pool; partial depletion leads to the formation of defective centrioles that recruit less than wild-type levels of PCM. | Kirkham, 2003; Leidel, 2003 |
| SAS-5 | sas-5 (F35B12.5) | YES | ? | Centriole protein required for centriole assembly; physically interacts with SAS-6; centriolar SAS-5 shuttles continuously between centrioles and the cytoplasm; requires ZYG-1 and SAS-6, but not SAS-4 to localize to centrioles; required for the localization of SAS-4 and SAS-6 to centrioles. Like SAS-4, partial depletion of SAS-5 leads to the formation of defective centrioles that recruit less than wild-type levels of PCM. | Delattre, 2004; Dammermann, 2004; Leidel, 2005 |
| SAS-6 | sas-6 (Y45F10D.9) | YES | HsSAS-6; DKFZP761 A078 | Centriole protein required for centriole assembly; like SAS-4, is recruited to centrioles once per cell cycle and does not subsequently exchange; physically interacts with SAS-5; requires ZYG-1 and SAS-5, but not SAS-4 to be recruited to centrioles; required for the localization of SAS-4 and SAS-5. Like SAS-4, partial depletion of SAS-5 leads to the formation of defective centrioles that recruit less than wild-type levels of PCM. The human homolog of SAS-6 localizes to centrioles and is required for centriole duplication. | Dammermann, 2004; Leidel, 2005 |
| ZYG-1 | zyg-1 (F59E12.2) | YES | ? | Centriole protein required for centriole assembly; localizes to centrioles throughout the cell cycle, but in contrast to SAS-4-6 is not detected on centrioles in sperm; required to target SAS-4, SAS-5 and SAS-6 to centrioles; atypical protein kinase that autophosphorylates in vitro, but in vivo substrates remain unknown; zyg-1 mutation interacts genetically with a mutation in spd-2. | O'Connell, 2001; Kemp, 2004; Delattre, 2004; Dammermann, 2004 |
| SPD-2 | spd-2 (F32H2.3) | YES | FLJ10352 | Bi-functional protein required for new centriole formation and for the assembly of the PCM around the centrioles; localizes to both centrioles and the PCM; spd-2 mutation interacts genetically with mutations in both zyg-1 and spd-5; the SPD-2 domain shares homology to the ASP protein family. | O'Connell, 2000; Kemp, 2004; Pelletier, 2004 |
| SPD-5 | spd-5 (F56A3.4) | YES | PCM component required for PCM assembly; required to recruit all tested PCM proteins; in SPD-5 depleted embryos centrosomal microtubule asters are absent and spindle assembly fails; depleted embryos also exhibit a severe defect in new centriole assembly, possibly due to failure to recruit gamma-tubulin to promote the formation of centriolar microtubules; a spd-5 mutation interacts genetically with mutations in spd-2 and dynein heavy chain. | Hamill, 2002; Kemp, 2004; Dammermann, 2004 | |
| AIR-1 | air-1 (K07C11.2) | YES | Aurora A | PCM component required for PCM assembly; localization appears more peripheral than that of gamma-tubulin, extending out along centrosomal microtubules; required to recruit additional PCM during centrosome maturation; in depleted embryos centrosomes remain small and spindle assembly fails. | Schumacher, 1998; Hannak, 2001 |
| gamma- tubulin | tbg-1 (F58A4.8) | YES | gamma- tubulin | Normal levels of SPD-5 are observed in depleted embryos indicating that it is not required for PCM formation; in depleted embryos, centrosomal microtubule asters fail to form during interphase, but robust asters assemble as embryos enter mitosis; although unknown mechanisms support partial assembly of mitotic centrosomal asters, the rate of centrosomal microtubule nucleation is severely compromised in depleted embryos indicating that the kinetically dominant pathway for the nucleation of centrosomal microtubules is gamma-tubulin dependent; depleted embryos also exhibit a severe defect in new centriole assembly. | Bobbinec, 2000; Strome, 2001; Hannak, 2002 |
| CeGrip-1 | gip-1 (H04J21.3) | YES | GCP3 | PCM component; orthologues form a heterotrimeric complex with gamma-tubulin in Xenopus, Drosophila, and S. cerevisiae; depletion prevents recruitment of gamma-tubulin to centrosomes; depletion phenotype is essentially identical to that observed in gamma tubulin depleted embryos. | Hannak, 2002 |
| CeGrip-2 | gip-2 (C45G3.3) | YES | GCP2 | PCM component; orthologues form a heterotrimeric complex with gamma-tubulin in Xenopus, Drosophila, and S. cerevisiae; depletion prevents recruitment of gamma-tubulin to centrosomes; depletion phenotype is essentially identical to that observed in gamma tubulin depleted embryos. | Hannak, 2002 |
| ZYG-9 | zyg-9 (F22B5.7) | YES | XMAP-215 | PCM component; physically interacts with TAC-1; this interaction is required for the efficient localization of the ZYG-9/TAC-1 complex to centrosomes; important regulator of microtubule dynamics; depletion phenotype is similar to that resulting from treatment of embryos with low doses of nocodozole which destabilize microtubules, pronuclear migration fails and a short spindle forms in the embryo posterior. | Matthews, 1998; Le Bot, 2003; Srayko, 2003; Bellanger, 2003 |
| TAC-1 | tac-1 (Y54E2A.3) | YES | TACC-1, 2, 3 | PCM component; physically interacts with ZYG-9; TAC-1 and ZYG-9 are dependent on each another for their localization to the centrosome. Depletion phenotype is similar to that resulting from treatment of embryos with low doses of nocodozole which destabilize microtubules, pronuclear migration fails and a short spindle forms in the embryo posterior. | Le Bot, 2003; Srayko, 2003; Bellanger, 2003 |
Several PCM proteins are required for the activity rather than the assembly of the PCM. The microtubule nucleating activity of the pericentriolar material requires the centrosomal tubulin isoform gamma-tubulin, and two gamma-tubulin associated proteins, CeGrip-1 and CeGrip-2. Embryos depleted of any of these proteins fail to form centrosomal microtubule asters during interphase (Hannak et al., 2002). As depleted embryos enter mitosis, relatively robust microtubule asters form around the centrosomes, suggesting that a gamma-tubulin independent pathway contributes to their assembly (Strome et al., 2001). However, chill and rewarm experiments reveal that the rate of centrosomal microtubule nucleation is highly compromised in gamma-tubulin depleted embryos and spindle assembly fails (Hannak et al., 2002; Strome et al., 2001). ZYG-9 and TAC-1 are two conserved PCM proteins that associate to form a complex that promotes microtubule growth. Embryos depleted of either protein exhibit defects in pronuclear migration, presumably because centrosomal microtubules are too short to capture the oocyte pronucleus, and small spindles form around the sperm chromatin in the embryo posterior (Bellanger and Gönczy, 2003; Le Bot et al., 2003; Matthews et al., 1998; Srayko et al., 2003).
Seven C. elegans proteins have been shown to contribute to new centriole assembly. Four of these, SAS-4 (Kirkham et al., 2003; Leidel and Gönczy, 2003), SAS-5 (Dammermann et al., 2004; Delattre et al., 2004; Schmutz and Spang, 2005), SAS-6 (Dammermann et al., 2004; Leidel et al., 2005), and the atypical protein kinase ZYG-1 (O'Connell et al., 2001), localize to centrioles and are specifically required for centriole formation. Depletion of any of these proteins by RNAi results in specific failure of centrosome duplication. In this characteristic phenotype (O'Connell et al., 2001), the two sperm centrioles separate after fertilization (as in wild-type) and organize the two centrosomes that form the poles of an apparently normal spindle during the first mitotic division. However, since daughter centrioles fail to form adjacent to each of the sperm centrioles, only one centriole instead of the normal two are released into each daughter cell when the centrosomes break down in telophase. Since each daughter cell contains only one centriole, it can form only one centrosome, and monopolar spindles are observed in both cells at the two-cell stage. SPD-2 is a bi-functional protein that localizes to centrioles as well as the PCM and is required for centriole assembly in addition to its role in PCM recruitment (Kemp et al., 2004; Pelletier et al., 2004). Embryos depleted of two pericentriolar material proteins, SPD-5 and gamma-tubulin, also exhibit severe defects in centriole assembly, leading to the idea that the PCM contributes to centriole assembly by recruiting gamma-tubulin, which may promote the assembly of the microtubules that make up the centriolar cylinders, as well as those nucleated by the PCM (Dammermann et al., 2004).
The formation of mitotic chromosomes begins when cohesin is loaded onto chromosomes and establishes cohesion between the duplicated chromosomes (sister chromatids) during DNA replication in S phase. Proteolytic cleavage of one of the cohesin subunits later in the cell cycle is thought to release the linkage between the sister chromatids to allow their segregation during anaphase (Haering and Nasmyth, 2003). The components of the C. elegans cohesin complex have been identified and depletion has revealed roles in mitotic chromosome segregation and the pairing of homologous chromosomes during meiosis (Table 3; Chan et al., 2003; Mito et al., 2003; Pasierbek et al., 2003).
Table 3. Mitotic chromosome proteins
| C. elegans protein | C. elegans gene | Required for embryonic viability? | Vertebrate orthologue | Summary of localization and functional analysis | Selected references |
|---|---|---|---|---|---|
| HIM-1 | him-1; smc-1; (F28B3.7) | YES | SMC-1 | Component of the cohesin complex; concentrated in the nucleus during interphase; by the metaphase to anaphase transition levels in the vicinity of the chromatin are reduced and protein appears excluded from the compacted chromatin. | Chan, 2003 |
| SMC-3 | smc-3; (Y47D3A.26) | YES | SMC-3 | Component of the cohesin complex; concentrated in the nucleus during interphase; by the metaphase to anaphase transition levels in the vicinity of the chromatin are reduced and protein appears excluded from the compacted chromatin. | Chan, 2003 |
| SCC-1 | scc-1; (F10G7.4) | YES | SCC-1 | Component of the cohesin complex; concentrated in the nucleus during interphase; by the metaphase to anaphase transition levels in the vicinity of the chromatin are reduced and protein appears to be excluded from the compacted chromatin; depletion results in a defect in mitotic chromosome segregation; the severity of the chromosome segregation defects is enhanced by simultaneous RNAi of him-1. | Chan, 2003; Mito, 2003 |
| SCC-3 | scc-3; (F18E2.3) | YES | SCC-3 | Component of the cohesin complex; concentrated in the nucleus during interphase; by the metaphase to anaphase transition levels in the vicinity of the chromatin are reduced and protein appears to be excluded from the compacted chromatin; depletion results in defects in chromosome segregation during mitosis and homologous pairing during meiosis. | Chan, 2003; Pasierbek, 2003; Moore, 2005 |
| TIM-1 | tim-1; csg-5; (Y75B8A.22) | YES | mTim1 | Component of the cohesin complex; concentrated in the nucleus during interphase; by the metaphase to anaphase transition levels in the vicinity of the chromatin are reduced and protein appears excluded from the compacted chromatin; RNAi of tim-1 in conjunction with him-1 results in more severe defects in chromosome segregation than either alone; required to recruit non-SMC cohesin subunits to chromatin before or during pre-meiotic S phase and to stabilize homologous chromosome associations during synapsis and sister chromatid cohesion in diplotene/ diakinesis. | Chan, 2003 |
| MIX-1 | mix-1; (M106.1) | YES | SMC-2 | Component of the condensin complex; required for mitotic chromosome condensation; chromosomal localization does not require kinetochore assembly; depletion results in severe chromatin bridging during anaphase; required for X-chromosome dosage compensation (repression). | Lieb, 1998; Hagstrom, 2002; Kaitna, 2002; Chan, 2004 |
| SMC-4 | smc-4; (F35G12.8) | YES | SMC-4 | Component of the condensin complex; required for mitotic chromosome condensation; chromosomal localization does not require kinetochore assembly; depletion leads to severe chromatin bridging during anaphase. | Hagstrom, 2002 |
| HCP-6 | hcp-6; (Y110A7A.1) | YES | CAP-D3, Heat (IIA) | Component of the condensin complex; required for mitotic chromosome condensation; chromosomal localization requires kinetochore assembly (fails to localize in CeCENP-C RNAi); required for mitotic chromosome condensation; depletion leads to severe chromatin bridging during anaphase. | Stear and Roth, 2002; Chan, 2004 |
| F55C5.4 | YES | CAP-G2; Heat (IIB) | Predicted component of the condensin complex based on sequence homology; no published localization or detailed depletion data. | Ono, 2003 | |
| KLE-2 | C29E4.2 | YES | CAP-H2; Kleisin γ (IIC) | Component of the condensin complex; required for mitotic chromosome condensation; depletion leads to severe chromatin bridging during anaphase. | Schleiffer, 2003 |
| AIR-2 | air-2; let-603; stu-7; cyk-6; (B0207.4) | YES | Aurora B | Aurora/Ip11- related serine/threonine protein kinase; chromosomal passenger protein, localizes to chromosomes between prophase and telophase and to microtubule bundles in the spindle midzone between anaphase and telophase; forms a complex with BIR-1, ICP-1, and CSC-1 and requires all three to localize to chromosomes; required for histone H3 phosphorylation, chromosome alignment and segregation, formation of the spindle midzone and cytokinesis. | Schumacher, 1998; Hsu et al., 2000; Kaitna, 2000; Severson, 2000; Oegema, 2001; Rogers 2002; Kaitna, 2002; Romano, 2003 |
| ICP-1, CeINCENP | icp-1; (Y39G10A R.13) | YES | INCENP | Chromosomal passenger protein, localizes to chromosomes between prophase and telophase and microtubule bundles in the spindle midzone between anaphase and telophase; required for chromosome alignment and segregation, formation of the spindle midzone and cytokinesis; chromosomal localization requires BIR-1 and CSC-1, but not AIR-2. Kinetochore assembly does not require ICP-1 and inhibiting kinetochore assembly by depletion of CENP-A (HCP-3) does not block the recruitment of ICP-1 to chromosomes. | Kaitna, 2000; Kaitna, 2002; Oegema, 2001; Romano, 2003 |
| BIR-1 | bir-1; (T27F2.3) | YES | Survivin | Chromosomal passenger protein, localizes to chromosomes between prophase and telophase and microtubule bundles in the spindle midzone between anaphase and telophase; chromosomal localization requires CSC-1 and ICP-1, but not AIR-2; required for chromosome alignment and segregation, formation of the spindle midzone and cytokinesis. | Fraser, 1999; Kaitna, 2000; Speliotes, 2000; Oegema, 2001; Romano, 2003 |
| KLP-19 | klp-19 | YES | kinesin-4; Kif4 | Localizes to the chromatin between the kinetochores; depletion results in aberrant chromosome motions during prometaphase, chromosome misalignment and the formation of multiple chromatin bridges during anaphase; thought to reduce the frequency with which individual diffuse kinetochores become attached to microtubules emanating from both spindle poles (merotelic attachment) by interacting with microtubules to stabilize an orientation in which the two kinetochores directly face the spindle poles. | Powers, 2004 |
Two additional protein complexes, condensin and the chromosomal passenger complex, also have critical roles in the formation and segregation of mitotic chromosomes. In contrast to vertebrates, which have two complexes, condensins I and II, that mediate spatially distinct aspects of condensation (Hirano, 2004), C. elegans has only condensin II (Table 3, Hagstrom et al., 2002; Ono et al., 2003). Chromosome condensation is delayed in condensin depleted embryos, but DNA compaction is ultimately achieved, indicating the existence of condensin-independent mechanisms that can compact mitotic chromatin. However, sister chromatid pairs formed in the absence of condensin are structurally defective and cannot be efficiently separated from each other by the mitotic spindle (Hagstrom et al., 2002; Kaitna et al., 2002; Chan et al., 2004). Concurrent with, but largely independent of condensation, kinetochores assemble to create chromosomal attachment sites for spindle microtubules (described in greater detail below). Like condensin, the chromosomal passenger protein complex (including the aurora B kinase, AIR-2, BIR-1, ICP-1 and CSC-1) is recruited to mitotic chromosomes as they form and is required for their proper segregation, (Schumacher et al., 1998; Severson et al., 2000; Kaitna, 2000; Oegema, 2001; Kaitna et al., 2002; Rogers 2002; Romano 2004; Hsu et al., 2000). Although it does not appear to be required for kinetochore assembly in C. elegans, the passenger complex plays important roles in chromosome structure and likely functions to correct aberrant kinetochore-microtubule attachments (reviewed in Vagnarelli and Earnshaw, 2004).
Eukaryotes can be divided into two groups based on the architecture of their mitotic chromosomes. Monocentric organisms assemble kinetochores on a single localized chromosomal site defined by the presence of dedicated centromeric chromatin. In contrast, holocentric organisms, including C. elegans, assemble diffuse kinetochores along the entire poleward face of each sister chromatid (Figure 6A). Holocentric chromosome architecture is present in widely divergent and highly successful metazoan lineages (including nematodes, hemipteran insects, and lower plants) that constitute a large part of the earth’s biomass, and C. elegans has emerged as an important model system for studying holocentric chromosome architecture (reviewed in Dernburg, 2001; Maddox et al., 2004).
Despite differences in the extent of the chromosomal length occupied by the kinetochore, the molecular composition of kinetochores in C. elegans and monocentric organisms is very similar (Table 4). Importantly, mitotic kinetochores in both monocentric and holocentric organisms assemble on a base of specialized centromeric chromatin defined by the presence of nucleosomes containing the histone H3 variant CENP-A (Buchwitz et al., 1999; Sullivan, 2001). Depletion of the C. elegans homolog of CENP-A leads to a characteristic “kinetochore-null” phenotype in which chromosomes fail to distribute over the spindle equator and to segregate (Oegema et al., 2001). This defect results from the failure to assemble kinetochores that can interact with spindle microtubules. To date only 4 proteins have been identified whose depletion gives a kinetochore-null defect: CENP-AHCP-3, CENP-CHCP-4, KNL-3 and KNL-1, which can be placed in a linear assembly hierarchy (Figure 6B) with CENP-AHCP-3 at the top (Moore and Roth, 2001; Oegema et al., 2001; Desai et al., 2003; Cheeseman et al., 2004). KNL-3 and KNL-1 are components of a 10-protein complex that is critical to forming the outer domains of the kinetochore that interact with spindle microtubules (Figure 6B; Cheeseman et al., 2004; Desai et al., 2003). A number of other kinetochore proteins have been identified whose depletion results in less severe chromosome segregation defects (for examples see Howe et al., 2001; Moore et al., 1999; for a detailed summary see Table 4). The reproducible phenotypes obtained following protein depletion have been useful in grouping proteins that function together in the context of the kinetochore (Figure 6B; Cheeseman et al., 2004).
|
Figure 6. Mitotic kinetochores in the C. elegans embryo. (A) Images comparing the localized kinetochores in a vertebrate tissue culture cell with monocentric chromosomes to the diffuse kinetochores in a C. elegans embryo with holocentric chromosomes. (B) Schematic illustrating the hierarchy for mitotic kinetochore assembly based on pair-wise depletion and localization assays, phenotypic analysis, and biochemical purifications. Dotted lines indicate groups of proteins that have been shown to co-purify from C. elegans extracts in immunoprecipitations and tagged protein isolations. The colored ovals group proteins together whose individual depletions result in a similar phenotype. (Red) “Kinetochore Null”/KNL proteins whose depletion is characterized by a failure to assemble kinetochores that are competent to form spindle microtubule attachments. Consequently, segregation is severely defective and spindle poles separate prematurely. (Blue) “MIS” proteins whose depletion results in relatively subtle chromosome segregation defects. In MIS embryos, kinetochore assembly occurs but at a slower rate and to a reduced extent relative to wild-type. (Yellow) “NDC” proteins whose depletion results in a chromosome alignment and segregation defects of intermediate severity, relative to the KNL and MIS classes. In NDC embryos, attachments that can sustain tension fail to form. Consequently, spindle poles separate prematurely. (Green) HCP/CLASP proteins, whose depletion causes sister chromatids to co-segregate to the same spindle pole. This defect likely arises from an inability to polymerize microtubules at kinetochores. (C) Schematic illustrating the temporal window during the first mitotic division when each of the indicated proteins localizes to kinetochores. Figure and images courtesy of Susan Kline-Smith and Arshad Desai, respectively.
Table 4. Kinetochore proteins
| C. elegans protein | C. elegans gene | Required for embryonic viability? | Vertebrate orthologue | Summary of localization and functional analysis | Selected references |
|---|---|---|---|---|---|
| CeCENP-A, HCP-3 | hcp-3; (F58A4.3) | YES | CENP-A | Histone H3-variant; thought to localize to chromatin throughout the cell cycle; found in nuclear foci during interphase and concentrated on the chromatin in a stripe that runs along the poleward face of each sister chromatid during mitosis; CeCENP-A is required for the localization of all other kinetochore components that have been tested; depletion results in failure to align and segregate mitotic chromosomes and premature spindle elongation due to the inability of the kinetochores to attach to spindle microtubules; a highly homologous gene (F54C8.2) is likely also targeted by dsRNAs generated against CeCENP-A. The relative abundance of CeCENP-A versus F54C8.2 is not known. C. briggsae does not have 2 genes encoding CENP-A like proteins, suggesting that F54C8.2 may have arisen from a recent duplication. | Buchwitz, 1999; Oegema, 2001 |
| CeCENP-C, HCP-4 | hcp-4; (T03F1.9) | YES | CENP-C | Kinetochore localization (prophase to telophase) requires CeCENP-A; CeCENP-C is required for the localization of all kinetochore proteins except CeCENP-A; associates with the KNL-1/3 complex and may act as an adaptor to connect centromeric chromatin to the outer kinetochore; plays a role in the resolution of CeCENP-A chromatin into two paired “lines” on the replicated chromosome; depletion results in failure to align and segregate mitotic chromosomes and premature spindle elongation. | Moore, 2001; Oegema, 2001; Desai, 2003; Cheeseman, 2004 |
| KNL-3 | knl-3 (T10B5.6) | YES | ? | Kinetochore localization (prophase to telophase) requires CeCENP-A and CeCENP-C; KNL-3 levels at the kinetochore are reduced in embryos depleted of MIS-12, KBP-1, KBP-2 and KNL-1; depletion results in failure to align and segregate mitotic chromosomes and premature spindle elongation; component of the 10-protein KNL-1/3 complex required to assemble an outer kinetochore that can make microtubule attachments. | Cheeseman, 2004 |
| KNL-1 | knl-1 (C02F5.1) | YES | AF15q14 | Kinetochore localization (prophase to telophase) requires CeCENP-A, CeCENP-C and KNL-3; KNL-1 levels at the kinetochores are reduced in embryos depleted of NDC-80 and HIM-10; depletion results in failure to align and segregate mitotic chromosomes and premature spindle elongation. Component of the 10-protein KNL-1/3 complex required to assemble an outer kinetochore that can make microtubule attachments. | Desai, 2003 Cheeseman, 2004 |
| MIS-12 | mis-12 (Y47G6A.24) | YES | Mis12 | Kinetochore localization (prophase to telophase) requires KNL-3 but not KNL-1; relatively weak chromosome missegregation phenotype seen in depleted embryos consistent with a delay in the formation of chromosome- spindle attachments; identified by sequence homology to human and fission yeast Mis12 and as a component of the KNL-1/3 complex. | Cheeseman, 2004 |
| KBP-1 | kbp-1 (R13F6.1) | YES | ? | Localizes to kinetochores from prophase to telophase; relatively weak chromosome missegregation phenotype seen in depleted embryos consistent with a delay in the formation of chromosome- spindle attachments; identified as a component of the KNL-1/3 complex. | Cheeseman, 2004 |
| KBP-2 | kbp-2 (F26F4.13) | YES | ? | Localizes to kinetochores from prophase to telophase; relatively weak chromosome missegregation phenotype seen in depleted embryos consistent with a delay in the formation of chromosome- spindle attachments; identified as a component of the KNL-1/3 complex. | Cheeseman, 2004 |
| NDC-80 | ndc-80 (W01B6.9.1) | YES | Ndc80/ HEC | Localizes to kinetochores from prophase to telophase; kinetochore localization requires KNL-1 and HIM-10 in addition to chromatin- proximal components; depletion results in chromosome missegregation and premature spindle elongation; phenotype is less severe than a kinetochore-null but more severe than that of MIS-12, suggests defect in the ability to form microtubule attachments that can withstand tension; member of the NDC-80 subcomplex, which is part of the larger KNL-1/3 complex. | Desai, 2003 Cheeseman, 2004 |
| HIM-10 Nuf2HIM-10 | him-10 (R12B2.4) | YES | Nuf2 | Localizes to kinetochores from prophase to telophase; kinetochore localization requires KNL-1 and NDC-80, in addition to chromatin- proximal components. Depletion disrupts kinetochore ultrastructure, resulting in chromosome missegregation and premature spindle elongation; phenotype is less severe than a kinetochore-null but more severe than that of MIS-12, suggests defect in the ability to form microtubule attachments that can withstand tension; member of the NDC-80 subcomplex, which is part of the larger KNL-1/3 complex. | Howe, 2001 Desai, 2003 Cheeseman, 2004 |
| KBP-3 | kbp-3 (F26H11.1) | YES | Spc25 | Localizes to kinetochores from prophase to telophase; depletion results in chromosome missegregation and premature spindle elongation; phenotype is less severe than a kinetochore-null but more severe than that of MIS-12, suggests defect in the ability to form microtubule attachments that can withstand tension; member of the NDC-80 subcomplex, which is part of the larger KNL-1/3 complex. | Cheeseman, 2004 |
| KBP-4 | kbp-4 (Y92C3B.1) | YES | ? | Localizes to kinetochores from prophase to telophase. Depletion results in chromosome missegregation and premature spindle elongation; phenotype is less severe than a kinetochore-null but more severe than that of MIS-12, suggests defect in the ability to form microtubule attachments that can withstand tension; identified as a KNL-1/3-associated protein. | Cheeseman, 2004 |
| KBP-5 | kbp-5 (C34B2.2) | NO | ? | Localizes to kinetochores from prophase to telophase. Identified as a KNL-1/3-associated protein. | Cheeseman, 2004 |
| HCP-1 HCP-2 | hcp-1 (ZK1055.1) hcp-2 (T06E4.1) | YES* *When co-depleted | CENP-F? | Functionally redundant proteins that localize to the region of the mitotic spindle and to kinetochores between late prometaphase and early anaphase; kinetochore localization requires CeCENP-A/C, KNL-1, and BUB-1, but not the NDC-80 subcomplex; depletion perturbs chromosome alignment and segregation. HCP-1 and 2 physically associate with CLASPCLS-2. | Moore, 1999; Desai, 2003; Stear, 2004; Encalada, 2004; Cheeseman, 2005 |
| CLASPcls-2 (CLS-2) | cls-2 (R107.6) | YES | CLASP | Microtubule- associated protein that localizes to spindle poles, the region of the miotic spindle and kinetochores; prominent kinetochore localization is seen at metaphase. Kinetochore localization requires KNL-1 and HCP-1/2, but not the NDC-80 subcomplex. Depletion perturbs chromosome alignment and segregation. CLASPCLS-2 associates with HCP-1/2. | Desai, 2003; Cheeseman, 2005 |
| CeMCAK (KLP-7) | klp-7 (K11D9.1) | YES | MCAK | Kinesin-13 microtubule depolymerase that localizes to kinetochores between early prometaphase and telophase, and to spindle poles throughout mitosis; kinetochore localization requires CeCENP-A and CeCENP-C, but not KNL-1 or KNL-3; depletion results in snapping of the anaphase spindle, suggesting excessive astral pulling forces or a defect in spindle midzone formation/ stability. | Oegema, 2001; Grill, 2001; Desai, 2003; Powers, 2004 |
| LIS-1 | lis-1 (T03F6.5) | YES | LIS1 | Microtubule associated protein that localizes to the cell cortex, nuclear periphery, kinetochores, and microtubule asters; kinetochore localization requires CeCENP-C, but not dynein; depletion results in multiple defects in spindle positioning, pronuclear migration, and centrosome separation, leading to gross missegregation of chromosomes. | Cockell, 2004 |
| CZW-1 | czw-1 (F20D12.4) | YES | Zw10 | Predicted kinetochore protein based on sequence homology; partial depletion results in chromosome missegregation; penetrant depletion results in sterility | Starr, 1998 |
| ROD-1 | rod-1 (F55G1.4) | YES | Rod | Predicted kinetochore protein based on sequence homology; depletion results in chromosome missegregation. | Scaerou, 2001 |
| BUB-1 | bub-1 (R06C7.8) | YES | Bub1 | Mitotic checkpoint pathway serine/threonine protein kinase; kinetochore localization (prophase to metaphase) requires CeCENP-A, CeCENP-C, and KNL-1; depletion results in misalignment and missegregation of chromosomes. | Oegema, 2001; Desai, 2003; Encalada, 2004 |
| BUB-3 | Y54G9A.6 | NO | Bub3 | Predicted mitotic checkpoint pathway protein based on sequence homology. | |
| MDF-1 | mdf-1 (C50F4.11) | NO | Mad1 | Mitotic checkpoint pathway protein based on sequence homology; mutant worms fail to thrive due to accumulated chromosomal abnormalities; required to delay the embryonic cell cycle in response to spindle defects. | Kitagawa, 1999; Encalada, 2004 |
| MDF-2 | mdf-2 (Y69A2AR. 30) | NO | Mad2 | Mitotic checkpoint pathway protein with similar functions as MDF-1; depletion of MDF-2 also overrides the mitotic arrest induced by anoxia, suggesting that survival under very low oxygen levels is promoted by activation of the mitotic checkpoint. | Kitagawa, 1999; Nystul, 2003; Encalada, 2004 |
| SAN-1 | san-1 (ZC328.4) | NO | BubR1 | Mitotic checkpoint pathway protein that localizes to kinetochores after nuclear envelope breakdown. SAN-1 is the homologue of budding yeast MAD3 protein (their vertebrate orthologue, BubR1, has a serine/threonine kinase domain fused to the Mad3 homology region). SAN-1 was identified in a genetic screen for anoxia-sensitive mutants, highlighting a link between survival under very low oxygen levels and mitotic arrest mediated by the mitotic checkpoint pathway. | Nystul, 2004 |
In addition to its role in directing kinetochore assembly, CENP-C has also been implicated in sister kinetochore resolution, the process by which the kinetochores on the two sister chromatids resolve from one another, coming to rest on opposite sides of the mitotic chromosome (Moore and Roth, 2001). Defects in sister kinetochore resolution in CENP-C depleted embryos can be suppressed by RNAi or mutation of cohesin subunits suggesting that resolution of the sister kinetochores during the assembly of mitotic chromosomes is normally facilitated by loss of cohesion between the sister centromeres (Moore et al., 2005). In addition to kinetochore-localized proteins, the chromokinesin KLP-19, which localizes to the chromatin between the diffuse kinetochores, is also critical for chromosome segregation. Interactions between KLP-19 and spindle microtubules are postulated to generate a pushing force that rotates chromosomes to orient the sister kinetochores to face opposite spindle poles (Table 3; Powers et al., 2004), thereby reducing the possibility that a single kinetochore will become incorrectly attached to microtubules coming from both spindle poles.
In vertebrates and Drosophila, spindle assembly results from the superposition of two distinct mechanisms: (1) an inside out chromatin-based mechanism in which Ran-GTP produced in proximity to chromatin activates factors that promote the assembly and self-organization of microtubules to form a bipolar spindle, and (2) an outside in mechanism in which centrosomal microtubule asters reinforce spindle bipolarity and position the spindle within the cell (Gadde and Heald, 2004). In contrast, C. elegans appears to lack a significant inside out pathway and primarily utilizes an outside in centrosome-based mechanism to form its spindles. Consistent with this idea, Eg5, a mitotic kinesin that promotes anti-parallel microtubule sliding and plays an essential role in the chromatin-based pathway in vertebrates and Drosophila, is not required for spindle assembly in C. elegans (Bishop et al., 2005). Conversely, whereas centrosomes are not essential for spindle assembly in Drosophila or vertebrates (Khodjakov et al., 2000; Megraw et al., 1999; Rebollo et al., 2004; Vaizel-Ohayon and Schejter, 1999), spindles fail to form in C. elegans embryos that lack functional centrosomes (Hamill et al., 2002; Kemp et al., 2004; Pelletier et al., 2004) and the majority of characterized C. elegans proteins that are required for spindle assembly (Sõnnichsen et al., 2005) are known centrosome components. Interestingly, essentially perfect “half spindles” form in embryos that have a single centrosome instead of the normal two (Kirkham et al., 2003; O'Connell et al., 2001), indicating that the mechanisms that regulate spindle length in the C. elegans embryo are largely independent of bipolarity. Experiments in which the size of mitotic centrosomes is modulated by varying the levels of centriole components have also revealed an interesting positive correlation between half-spindle length and centrosome size (Delattre et al., 2004; Kirkham et al., 2003).
Concomitant with nuclear envelope break down, centrosomal microtubules penetrate the nuclear space and begin to interact with chromosomes. Spindle assembly is completed in the subsequent ~2.5 minute period between nuclear envelope breakdown and anaphase onset (see Figure 2). Tomographic reconstruction of a C.elegans mitotic spindle from electron micrographs indicates that C. elegans mitotic spindles consist primarily of microtubules that connect centrosomes to kinetochores. These kinetochore microtubules do not form bundles that resemble the kinetochore fibers present in vertebrate and Drosophila cells, probably because of the holocentric nature of C. elegans chromosomes (O'Toole et al., 2003). Assembly of a stable mitotic spindle requires kinetochores that can form stable bipolar microtubule attachments. In the absence of functional kinetochores, the two spindle poles are abruptly pulled apart at a time that corresponds to the onset of cortical pulling forces on the two centrosomes that normally asymmetrically position the spindle within the embryo (Oegema et al., 2001). The abrupt separation of the spindle poles in the absence of functional kinetochores is similar to that brought about by severing the spindle with a UV microbeam (Grill et al., 2001), confirming the critical role of kinetochores in maintaining spindle integrity.
Chromosome segregation typically consists of two components: (1) anaphase A, in which the chromosomes move towards the spindle poles and, (2) anaphase B in which the spindle poles separate from each other with the chromosomes in tow. In C. elegans the majority of chromosome movement is due to anaphase B (Oegema et al., 2001), powered by a combination of cortical forces that pull on the centrosomal microtubule asters to separate them, and pushing forces generated by the array of overlapping microtubules, called the central spindle, that forms between the separating chromosomes. If the mitotic spindle is severed during anaphase using a UV micro beam, the two spindle poles separate with increased velocity (Grill et al., 2001). This experiment demonstrates the existence of pulling forces during the first mitotic division and suggests that the central spindle normally acts to limit the rate of pole separation. However, in embryos in which G protein signaling that mediates cortical pulling forces is disrupted, spindles still elongate (Colombo et al., 2003) albeit at a slower rate, indicating that other spindle intrinsic forces contribute to anaphase spindle elongation.
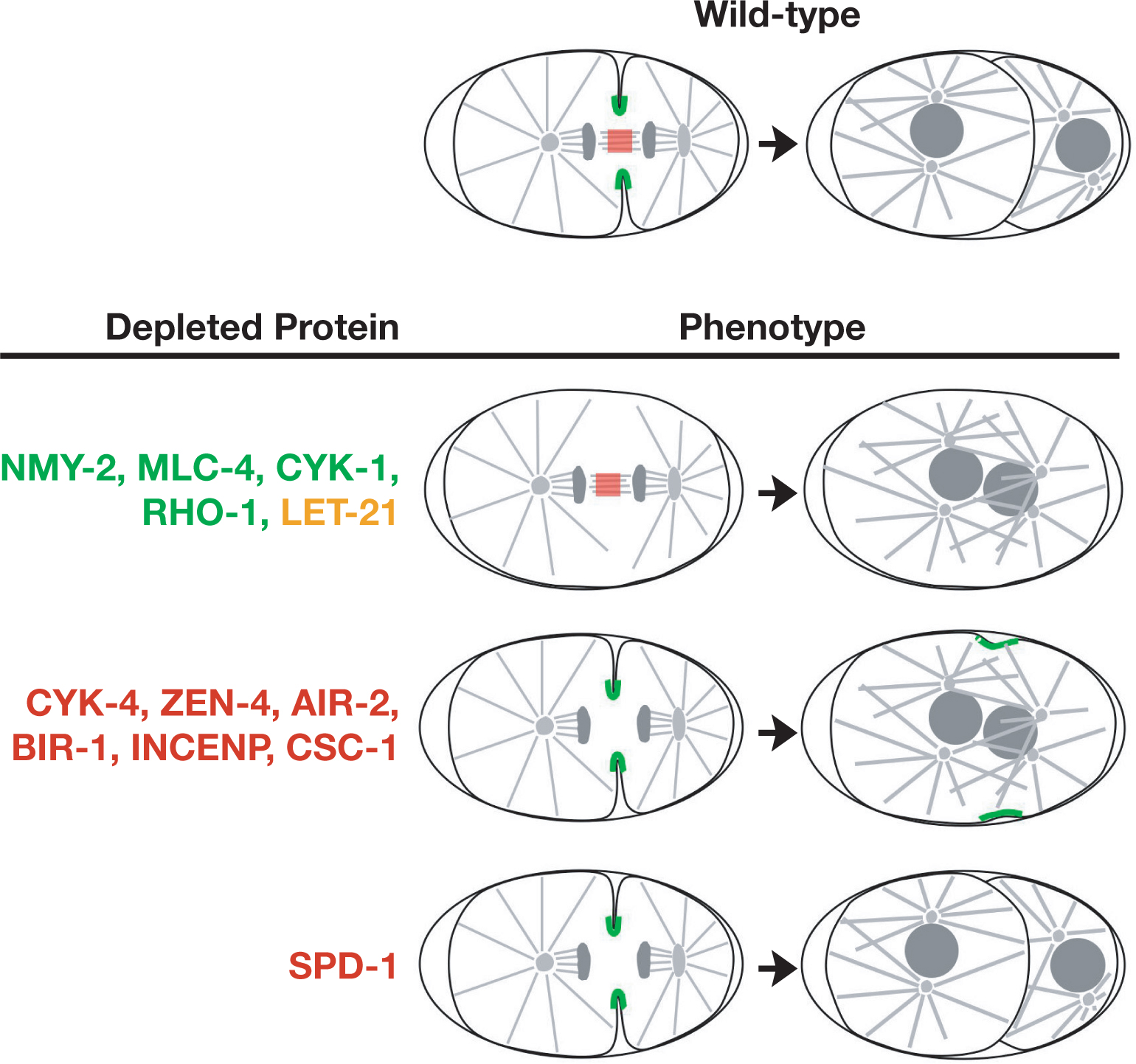
During cytokinesis, signals from the anaphase spindle trigger the assembly of an equatorial cortical contractile ring enriched in actin and myosin II. Constriction of the contractile ring changes the shape of the cell to facilitate cell division (reviewed in Glotzer, 2005). The components of the microtubule and actin cytoskeletons that are known to contribute to cytokinesis in the C. elegans embryo can be partitioned into three classes (Table 5). The first class consists of proteins required for cortical acto-myosin contractility (Figure 7). Embryos depleted of proteins in this class have defects in all cortical contractile events including: polar body extrusion, anterior cortical ruffling, pseudocleavage, and the initiation of a furrow during cytokinesis. This class includes the small GTPase RHO-1 (Jantsch-Plunger et al., 2000) and its activating GEF (LET-21; Dechant and Glotzer, 2003), which are thought to be at the top of the signaling cascade that regulates cortical contractility (Glotzer, 2005). Downstream are proteins that regulate the assembly of the contractile ring and its constriction in response to activated Rho. CYK-1 (Severson et al., 2002; Swan et al., 1998), a member of the formin family of proteins, is a Rho effector that promotes the assembly of actin filaments. The assembly and contractility of non-muscle myosin II (NMY-2; Guo and Kemphues, 1996), the motor that powers acto-myosin contractility, is regulated by phosphorylation of its regulatory light chain, MLC-4 (Shelton et al., 1999). Phosphorylation of MLC-4 activates myosin II, allowing it to form bi-polar filaments that can interact with actin to generate cortical ingression (Citi and Kendrick-Jones, 1987). One of the Rho effectors that regulates MLC-4 phosphorylation during cytokinesis is the Rho kinase homolog LET-502 (for details see Table 5; Piekny and Mains, 2002).
Table 5. Cytoskeletal proteins required for cytokinesis
| C. elegans protein | C. elegans gene | Required for embryonic viability? | Vertebrate orthologue | Summary of localization and functional analysis | Selected references |
|---|---|---|---|---|---|
| NMY-2 | YES | non-muscle myosin II | Localizes to the contractile ring during cytokinesis as well as to other cortical contractile structures; motor that powers contraction by moving towards the barbed end of actin filaments; all cortical contractile events fail in depleted embryos including polar body formation, ruffling, and cleavage furrow ingression. NMY-2 is also required for the establishment of polarity. | Guo, 1996; Munro, 2004 | |
| MLC-4 | mlc-4 (C56G7.1) | YES | non-muscle myosin II regulatory light chain | Regulates the ability of myosin II to form filaments and interact with actin; localizes to the cortical contractile ring during cytokinesis as well as to other cortical contractile structures; activated by phosphorylation; all cortical contractile events fail in depleted embryos including polar body formation, ruffling, and cleavage furrow ingression; MLC-4 is also required for the establishment of polarity. | Shelton, 1999 |
| LET-502 | let-502 (C10H11.9) | YES | Rho-binding kinase (ROK) | Rho-binding serine/threonine kinase that localizes to the contractile ring; promotes myosin II contractility by increasing the phosphorylation of MLC-4; based on work in other systems, it is thought to do this by both directly phosphorylating MLC-4 and also by phosphorylating and inactivating myosin phosphatase; inhibition of Rho-kinase slows the rate of furrow ingression but does not prevent furrow assembly or ingression, suggesting the existence of redundant MLC-4 kinases; failed cell divisions occur apparently at random in let-502 embryos, with many divisions being normal. | Piekny, 2002 |
| MEL-11 | mel-11 (C06C3.1) | YES | Myosin phosphatase targeting subunit (MYPT) | Regulatory subunit of myosin phosphatase; inhibits cortical contraction by de-phosphory- lating the regulatory light chain of myosin II; mutations in mel-11 result in ectopic furrowing and faster furrow ingression; LET-502 and MEL-11 co-localize in cleavage furrows and their mutations alleviate one another’s effects. | Piekny, 2002 |
| RHO-1 | rho-1 (Y51H4A.3) | YES | RhoA | Small GTPase thought to connect signaling by the anaphase spindle to assembly and ingression of a cortical contractile ring; all cortical contractile events fail in depleted embryos including polar body formation, ruffling, and cleavage furrow ingression; has not been localized in C. elegans but active Rho has been localized to the contractile ring in other systems. | Jantsch-Plunger, 2000 |
| LET-21 | let-21 (T19E10.1) | YES | Ect2 | Guanine nucleotide exchange factor that activates RHO-1; all cortical contractile events fail in depleted embryos including polar body formation, ruffling, and cleavage furrow ingression; has not been localized in C. elegans, but in other systems has been localized to microtubule bundles in the central spindle and to the contractile ring. | Dechant, 2003 |
| CYK-1 | cyk-1 (F11H8.4) | YES | formins | A member of the formin family of proteins thought to promote actin assembly in response to activation of Rho family GTPases; localizes to the cleavage furrow and is required to initiate furrow ingression. | Swan, 1998; Severson, 2002 |
| PFN-1 | pfn-1 (Y18D10A. 20) | YES | profillin | One of three C. elegans homologs of the actin binding protein profilin; the only C. elegans profilin that is required in the early embryo; depleted embryos exhibit defects in furrow ingression and the establishment of polarity; thought to function together with CYK-1 to promote actin assembly in response to activation by Rho family GTPases. | Severson, 2002 |
| ANI-1 | ani-1 (Y49E10.19) | YES | Anillin | One of three C. elegans homologs of the actin-binding protein anillin; the only anillin homolog that appears to have a role in cortical events in the early embryo; localizes to the contractile ring, as well as to other cortical contractile structures, and is required for the septins to concentrate in the furrow; not essential for embryonic cytokinesis, but required for polar body extrusion, cortical ruffling, and pseudocleavage. | Maddox, 2005 |
| UNC-59; UNC-61 | unc-59 (W09C5.2); unc-61 (Y50E8A.4) | NO | septins | The only C. elegans homologs of the septins, small GTPases that form heteromeric complexes that polymerize to form filaments; UNC-59 and UNC-61 are interdependent for their localization to the cortex and for their localization to the contractile ring; require ANI-1 to become enriched in the contractile ring, but are not required for the targeting of ANI-1; neither is essential for embryonic cytokinesis. | Nguyen, 2000; Maddox, 2005 |
| UNC-60A | unc-60 (C38C3.5) | YES | cofilin | Promotes actin turnover; concentrates in the cleavage furrow during cytokinesis; depleted embryos frequently exhibit cytokinesis defects; the unc-60 gene generates two ADF/cofilins, UNC-60A and UNC-60B, by alternative splicing that have different functions. UNC-60B is expressed not it embryos, but in somatic/adult tissues. | Ono, 2003 |
| ZEN-4 | zen-4 (M03D4.1) | YES | kinesin-6 family member MKLP1 | Kinesin that physically associates with CYK-4, a GAP for Rho family GTPases, to form a two protein complex called Centralspindlin; localizes to microtubule bundles in the spindle midzone and midbody; required for assembly of the spindle midzone and for the completion of cytokinesis, but not to form a furrow or initiate ingression. | Raich, 1998; Powers, 1998; Severson, 2000; Mishima, 2002 |
| CYK-4 | cyk-4 (K08E3.6) | YES | MgcRacGAP | A GTPase activating protein (GAP), an inhibitor for Rho family GTPases; associates with ZEN-4 to form a protein complex called Centralspindlin; localizes to microtubule bundles in the central spindle and midbody; required for assembly of the central spindle and for the completion of cytokinesis, but not to form a furrow or initiate ingression. | Jantsch-Plunger, 2000; Mishima, 2002 |
| AIR-2 | air-2 let-603 stu-7 cyk-6 (B0207.4) | YES | Aurora B | Mitotic serine/threonine kinase; component of a four-protein chromosomal passenger protein complex that localizes to chromosomes during mitosis and to microtubule bundles in the central spindle and midbody during anaphase/ telophase; required for assembly of the central spindle/midbody and for the completion of cytokinesis, but not to form a furrow or initiate ingression; also required for chromosome segregation (see Table 3). | Schumacher, 1998; Kaitna, 2000; Severson, 2000 |
| ICP-1 CeINCENP | icp-1 (Y39G10AR. 13) | YES | INCENP | Component of a four-protein chromosomal passenger protein complex that localizes to chromosomes during mitosis and to microtubule bundles in the central spindle and midbody during anaphase/ telophase; phosphorylated by AIR-2 and stimulates AIR-2 kinase activity; required for assembly of the central spindle/midbody and for the completion of cytokinesis, but not to form a furrow or initiate ingression; also required for chromosome segregation (see Table 3). | Kaitna, 2000; Oegema, 2001; Bishop, 2002; Romano, 2004 |
| BIR-1 | bir-1 (T27F2.3) | YES | Survivin | Component of a four-protein chromosomal passenger protein complex that localizes to chromosomes during mitosis and to microtubule bundles in the central spindle and midbody during anaphase/ telophase; required for assembly of the central spindle/midbody and for the completion of cytokinesis, but not to form a furrow or initiate ingression; also required for chromosome segregation (see Table 3). | Fraser, 1999; Kaitna, 2000; Speliotes, 2000; Romano, 2004 |
| CSC-1 | csc-1 (Y48E1B.12) | YES | Borealin, Dasra A/B | Component of a four-protein chromosomal passenger protein complex that localizes to chromosomes during mitosis and to microtubule bundles in the central spindle and midbody during anaphase/ telophase; required for assembly of the central spindle/midbody and for the completion of cytokinesis, but not to form a furrow or initiate ingression; also required for chromosome segregation (see Table 3). | Romano, 2004 |
| SPD-1 | spd-1 (Y34D9A.4) | YES | PRC1 | Localizes to microtubule bundles in the spindle midzone and midbody and to nuclei, as well as to short segments along the length of astral microtubules; required to form microtubule bundles in the central spindle; not required for the cytokinesis up to the four-cell stage, but cytokinesis often fails in EMS in depleted embryos. | Verbrugghe, 2004 |
A second group of proteins is required for the formation of the spindle midzone, an array of microtubule bundles that forms between the separating chromosomes during anaphase (Glotzer, 2005). As cytokinesis progresses, the microtubule bundles in the central spindle compact to form a single structure called the midbody. The cleavage furrow constricts around the midbody, which is thought to promote membrane fusion to complete cytokinesis and generate the two topologically distinct daughter cells (reviewed in Glotzer, 2005). During cytokinesis in embryos depleted of proteins required for central spindle assembly, a furrow initiates between the two asters, but cytokinesis fails to complete because the midbody is absent, and the furrow ultimately regresses. Depleted embryos also have defects in polar body extrusion (highly asymmetric cytokineses that bisect the midbodies of the small meiosis I and II spindles), but are able to form cortical ruffles that concentrate in the embryo anterior during polarity establishment and a pseudocleavage furrow. The microtubule bundles in the spindle midzone are also thought to function early in cytokinesis to signal to the cortex to promote assembly of the contractile ring. Disruption of the spindle midzone does not prevent furrow formation because this midzone-based signal is redundant with signals from the microtubule asters. In embryos in which the astral signaling mechanism is blocked, the spindle midzone becomes essential for furrow ingression (Dechant and Glotzer, 2003). Proteins required to form the central spindle include: (1) a two protein complex called centralspindlin that includes the kinesin ZEN-4 and CYK-4, a GAP for Rho family GTPases (Jantsch-Plunger et al., 2000; Mishima et al., 2002; Powers et al., 1998; Raich et al., 1998; Severson et al., 2000), and (2) the 4 protein chromosomal passenger protein complex containing the C. elegans homolog of the aurora B kinase, AIR-2 (Bishop and Schumacher, 2002; Fraser et al., 1999; Kaitna et al., 2000; Oegema et al., 2001; Romano et al., 2003; Schumacher et al., 1998; Speliotes et al., 2000).
|
Figure 7. Proteins required for cytokinesis can grouped based on their depletion phenotypes. Proteins that localize to the cortical contractile ring in C. elegans are shown in green. Rho has not been localized in C. elegans but activated Rho localizes to the furrow in other systems. Proteins that localize to the spindle midzone/midbody in C. elegans are shown in red. LET-21 (yellow) has not been localized in C. elegans, but is reported to localize to both the contractile ring and the spindle midzone/midbody in other systems. In embryos depleted of LET-21 or the contractile ring components NMY-2, MLC-4, CYK-1 or RHO-1, a normal spindle midzone forms between the separating chromosomes but furrow formation is inhibited and ingression often fails. In embryos depleted of the spindle midzone/midbody components CYK-4 and ZEN-4 or the chromosomal passenger proteins AIR-2, BIR-1, INCENP and CSC-1, a normal spindle midzone fails to form. Cleavage furrows form and ingress, but cytokinesis fails to complete and the furrow ultimately regresses. In embryos depleted of the spindle midzone/midbody component, SPD-1, the first cytokinesis succeeds despite defects in the structure of the spindle midzone/midbody.
A final protein, currently in a class of its own is SPD-1 (Verbrugghe and White, 2004). Like the six proteins listed above, SPD-1 is also required for the formation of microtubule bundles in the spindle midzone. However, in SPD-1 depleted embryos the first cytokinesis does not fail (although later embryonic cytokineses often do). It is not yet clear if this result indicates that the midbody is less compromised in spd-1 embryos than it is in embryos depleted of or mutant for the other six proteins required for midbody formation—or if centralspindlin and the chromosomal passengers have a separate role in the completion of cytokinesis that is independent of their role in assembling the central spindle/midbody. Three additional proteins, dynamin (Thompson et al., 2002), RAB-11 (Skop et al., 2001) and syntaxin 4 (SYN-4; Jantsch-Plunger and Glotzer, 1999), have been implicated in membrane fusion during abscission, which generates the two topologically distinct daughter cells, however their precise roles in this process remain to be elucidated.
The authors would like to thank Arshad Desai for critical reading of the manuscript and Amy Shaub Maddox, Susan Kline-Smith, Vincent Galy, Carrie Cowan, Eva Hannak and Stephan Grill for their contributions to the figures. Paul Maddox, Laurence Pelletier, Alexander Dammermann, Vincent Galy and Susan Kline-Smith provided invaluable assistance in constructing and referencing the tables. K.O. is a Pew Scholar in the Biomedical Sciences and is supported by funding from the Ludwig Institute for Cancer Research.
Albertson, D.G. (1984). Formation of the first cleavage spindle in nematode embryos. Dev. Biol. 101, 61–72. Article
Albertson, D.G., and Thomson, J.N. (1993). Segregation of holocentric chromosomes at meiosis in the nematode, Caenorhabditis elegans. Chromosome Res. 1, 15–26. Abstract Article
Askjaer, P., Galy, V., Hannak, E., and Mattaj, I.W. (2002). Ran GTPase cycle and importins α and β are essential for spindle formation and nuclear envelope assembly in living Caenorhabditis elegans embryos. Mol. Biol. Cell 13, 4355–4370. Abstract Article
Bamba, C., Bobinnec, Y., Fukuda, M., and Nishida, E. (2002). The GTPase Ran regulates chromosome positioning and nuclear envelope assembly in vivo. Curr. Biol. 12, 503–507. Abstract Article
Bellanger, J.M., and Gonczy, P. (2003). TAC-1 and ZYG-9 form a complex that promotes microtubule assembly in C. elegans embryos. Curr. Biol. 13, 1488–1498. Abstract Article
Bishop, J.D., Han, Z., and Schumacher, J.M. (2005). The Caenorhabditis elegans Aurora B kinase AIR-2 phosphorylates and is required for the localization of a BimC kinesin to meiotic and mitotic spindles. Mol. Biol. Cell 16, 742–756. Abstract Article
Bishop, J.D., and Schumacher, J.M. (2002). Phosphorylation of the carboxyl terminus of inner centromere protein (INCENP) by the Aurora B Kinase stimulates Aurora B kinase activity. J. Biol. Chem. 277, 27577–27580. Abstract Article
Bobinnec, Y., Fukuda, M., and Nishida, E. (2000). Identification and characterization of Caenorhabditis elegans γ-tubulin in dividing cells and differentiated tissues. J. Cell Sci. 113(Pt 21), 3747–3759.
Bornens, M. (2002). Centrosome composition and microtubule anchoring mechanisms. Curr. Opin. Cell Biol. 14, 25–34. Abstract Article
Brauchle, M., Baumer, K., and Gönczy, P. (2003). Differential activation of the DNA replication checkpoint contributes to asynchrony of cell division in C. elegans embryos. Curr. Biol. 13, 819–827. Abstract Article
Buchwitz, B.J., Ahmad, K., Moore, L.L., Roth, M.B., and Henikoff, S. (1999). A histone-H3-like protein in C. elegans. Nature 401, 547–548. Abstract Article
Callaini, G., and Riparbelli, M.G. (1990). Centriole and centrosome cycle in the early Drosophila embryo. J. Cell Sci. 97(Pt 3), 539–543. Abstract
Chan, R.C., Chan, A., Jeon, M., Wu, T.F., Pasqualone, D., Rougvie, A.E., and Meyer, B.J. (2003). Chromosome cohesion is regulated by a clock gene paralogue TIM-1. Nature 423, 1002–1009. Abstract Article
Chan, R.C., Severson, A.F., and Meyer, B.J. (2004). Condensin restructures chromosomes in preparation for meiotic divisions. J. Cell Biol. 167, 613–625. Abstract Article
Cheeks, R.J., Canman, J.C., Gabriel, W.N., Meyer, N., Strome, S., and Goldstein, B. (2004). C. elegans PAR proteins function by mobilizing and stabilizing asymmetrically localized protein complexes. Curr. Biol. 14, 851–862. Abstract Article
Cheeseman, I.M., MacLeod, I., Yates, J.R., III, Oegema, K., and Desai, A. (2005). The CENP-F-like proteins HCP-1 and HCP-2 Target CLASP to Kinetochores to Regulate Microtubule Dynamics. Curr. Biol.
Cheeseman, I.M., Niessen, S., Anderson, S., Hyndman, F., Yates, J.R., III, Oegema, K., and Desai, A. (2004). A conserved protein network controls assembly of the outer kinetochore and its ability to sustain tension. Genes Dev. 18, 2255–2268. Abstract Article
Citi, S., and Kendrick-Jones, J. (1987). Regulation of non-muscle myosin structure and function. Bioessays 7, 155–159. Abstract Article
Clark-Maguire, S., and Mains, P.E. (1994). Localization of the mei-1 gene product of Caenorhaditis elegans, a meiotic-specific spindle component. J. Cell Biol. 126, 199–209. Abstract Article
Cockell, M.M., Baumer, K., and Gönczy, P. (2004). lis-1 is required for dynein-dependent cell division processes in C. elegans embryos. J. Cell Sci. 117, 4571–4582. Article
Cohen, M., Tzur, Y.B., Neufeld, E., Feinstein, N., Delannoy, M.R., Wilson, K.L., and Gruenbaum, Y. (2002). Transmission electron microscope studies of the nuclear envelope in Caenorhabditis elegans embryos. J. Struct. Biol. 140, 232–240. Abstract Article
Colombo, K., Grill, S.W., Kimple, R.J., Willard, F.S., Siderovski, D.P., and Gönczy, P. (2003). Translation of polarity cues into asymmetric spindle positioning in Caenorhabditis elegans embryos. Science 300, 1957–1961. Abstract Article
Cowan, C.R., and Hyman, A.A. (2004). Asymmetric cell division in C. elegans: cortical polarity and spindle positioning. Annu. Rev. Cell Dev. Biol. 20, 427–453. Abstract Article
Cowan, C.R., and Hyman, A.A. (2004). Centrosomes direct cell polarity independently of microtubule assembly in C. elegans embryos. Nature 431, 92–96. Abstract Article
Dammermann, A., Muller-Reichert, T., Pelletier, L., Habermann, B., Desai, A., and Oegema, K. (2004). Centriole assembly requires both centriolar and pericentriolar material proteins. Dev. Cell 7, 815–829. Abstract Article
Dechant, R., and Glotzer, M. (2003). Centrosome separation and central spindle assembly act in redundant pathways that regulate microtubule density and trigger cleavage furrow formation. Dev. Cell 4, 333–344. Abstract Article
Delattre, M., Leidel, S., Wani, K., Baumer, K., Bamat, J., Schnabel, H., Feichtinger, R., Schnabel, R., and Gönczy, P. (2004). Centriolar SAS-5 is required for centrosome duplication in C. elegans. Nat. Cell Biol. 6, 656–664. Abstract Article
Dernburg, A.F. (2001). Here, there, and everywhere: kinetochore function on holocentric chromosomes. J. Cell Biol. 153, F33–F38. Abstract Article
Desai, A., Rybina, S., Muller-Reichert, T., Shevchenko, A., Hyman, A., and Oegema, K. (2003). KNL-1 directs assembly of the microtubule-binding interface of the kinetochore in C. elegans. Genes Dev. 17, 2421–2435. Abstract Article
Dutcher, S.K. (2003). Long-lost relatives reappear: identification of new members of the tubulin superfamily. Curr. Opin. Microbiol. 6, 634–640. Abstract Article
Encalada, S.E., Martin, P.R., Phillips, J.B., Lyczak, R., Hamill, D.R., Swan, K.A., and Bowerman, B. (2000). DNA replication defects delay cell division and disrupt cell polarity in early Caenorhabditis elegans embryos. Dev. Biol. 228, 225–238. Abstract Article
Encalada, S.E., Willis, J., Lyczak, R., and Bowerman, B. (2004). A spindle checkpoint functions during mitosis in the early Caenorhabditis elegans embryo. Mol. Biol. Cell 16, 1056–1070. Abstract Article
Fernandez, A.G., Gunsalus, K.C., Huang, J., Chuang, L.S., Ying, N., Liang, H.L., Tang, C., Schetter, A.J., Zegar, C., Rual, J.F., et al. (2005). New genes with roles in the C. elegans embryo revealed using RNAi of ovary-enriched ORFeome clones. Genome Res. 15, 250–259. Abstract Article
Fraser, A.G., James, C., Evan, G.I., and Hengartner, M.O. (1999). Caenorhabditis elegans inhibitor of apoptosis protein (IAP) homologue BIR-1 plays a conserved role in cytokinesis. Curr. Biol. 9, 292–301. Abstract Article
Fraser, A.G., Kamath, R.S., Zipperlen, P., Martinez-Campos, M., Sohrmann, M., and Ahringer, J. (2000). Functional genomic analysis of C. elegans chromosome I by systematic RNA interference. Nature 408, 325–330. Abstract Article
Fridkin, A., Mills, E., Margalit, A., Neufeld, E., Lee, K.K., Feinstein, N., Cohen, M., Wilson, K.L., and Gruenbaum, Y. (2004). Matefin, a Caenorhabditis elegans germ line-specific SUN-domain nuclear membrane protein, is essential for early embryonic and germ cell development. Proc. Natl. Acad. Sci. USA 101, 6987–6992. Article
Gadde, S., and Heald, R. (2004). Mechanisms and molecules of the mitotic spindle. Curr. Biol. 14, R797–R805. Abstract Article
Galy, V., Mattaj, I.W., and Askjaer, P. (2003). Caenorhabditis elegans nucleoporins Nup93 and Nup205 determine the limit of nuclear pore complex size exclusion in vivo. Mol. Biol. Cell 14, 5104–5115. Abstract Article
Glotzer, M. (2005). The molecular requirements for cytokinesis. Science 307, 1735–1739. Abstract Article
Golden, A., Sadler, P.L., Wallenfang, M.R., Schumacher, J.M., Hamill, D.R., Bates, G., Bowerman, B., Seydoux, G., and Shakes, D.C. (2000). Metaphase to anaphase (mat) transition-defective mutants in Caenorhabditis elegans. J. Cell Biol. 151, 1469–1482. Abstract Article
Goldstein, B., and Hird, S.N. (1996). Specification of the anteroposterior axis in Caenorhabditis elegans. Development 122, 1467–1474. Abstract
Gönczy, P., Echeverri, C., Oegema, K., Coulson, A., Jones, S.J., Copley, R.R., Duperon, J., Oegema, J., Brehm, M., Cassin, E., et al. (2000). Functional genomic analysis of cell division in C. elegans using RNAi of genes on chromosome III. Nature 408, 331–336. Abstract Article
Gönczy, P., Pichler, S., Kirkham, M., and Hyman, A.A. (1999). Cytoplasmic dynein is required for distinct aspects of MTOC positioning, including centrosome separation, in the one cell stage Caenorhabditis elegans embryo. J. Cell Biol. 147, 135–150. Abstract Article
Gönczy, P., Schnabel, H., Kaletta, T., Amores, A.D., Hyman, T., and Schnabel, R. (1999). Dissection of cell division processes in the one cell stage Caenorhabditis elegans embryo by mutational analysis. J. Cell Biol. 144, 927–946. Abstract Article
Grill, S.W., Gönczy, P., Stelzer, E.H., and Hyman, A.A. (2001). Polarity controls forces governing asymmetric spindle positioning in the Caenorhabditis elegans embryo. Nature 409, 630–633. Abstract Article
Gruenbaum, Y., Lee, K.K., Liu, J., Cohen, M., and Wilson, K.L. (2002). The expression, lamin-dependent localization and RNAi depletion phenotype for emerin in C. elegans. J. Cell Sci. 115, 923–929. Abstract
Gruenbaum, Y., Margalit, A., Goldman, R.D., Shumaker, D.K., and Wilson, K.L. (2005). The nuclear lamina comes of age. Nat. Rev. Mol. Cell Biol. 6, 21–31. Abstract Article
Guo, S., and Kemphues, K.J. (1996). A non-muscle myosin required for embryonic polarity in Caenorhabditis elegans. Nature 382, 455–458. Abstract Article
Haering, C.H., and Nasmyth, K. (2003). Building and breaking bridges between sister chromatids. Bioessays 25, 1178–1191. Abstract Article
Hagstrom, K.A., Holmes, V.F., Cozzarelli, N.R., and Meyer, B.J. (2002). C. elegans condensin promotes mitotic chromosome architecture, centromere organization, and sister chromatid segregation during mitosis and meiosis. Genes Dev. 16, 729–742. Abstract Article
Hamill, D.R., Severson, A.F., Carter, J.C., and Bowerman, B. (2002). Centrosome maturation and mitotic spindle assembly in C. elegans require SPD-5, a protein with multiple coiled-coil domains. Dev. Cell 3, 673–684. Abstract Article
Hannak, E., Kirkham, M., Hyman, A.A., and Oegema, K. (2001). Aurora-A kinase is required for centrosome maturation in Caenorhabditis elegans. J. Cell Biol. 155, 1109–1116. Abstract Article
Hannak, E., Oegema, K., Kirkham, M., Gönczy, P., Habermann, B., and Hyman, A.A. (2002). The kinetically dominant assembly pathway for centrosomal asters in Caenorhabditis elegans is γ-tubulin dependent. J. Cell Biol. 157, 591–602. Abstract Article
Hirano, T. (2004). Chromosome shaping by two condensins. Cell Cycle 3, 26–28. Abstract
Hird, S.N., and White, J.G. (1993). Cortical and cytoplasmic flow polarity in early embryonic cells of Caenorhabditis elegans. J. Cell Biol. 121, 1343–1355. Abstract Article
Howe, M., McDonald, K.L., Albertson, D.G., and Meyer, B.J. (2001). HIM-10 is required for kinetochore structure and function on Caenorhabditis elegans holocentric chromosomes. J. Cell Biol. 153, 1227–1238. Abstract Article
Hsu, J.Y., Sun, Z.W., Li, X., Reuben, M., Tatchell, K., Bishop, D.K., Grushcow, J.M., Brame, C.J., Caldwell, J.A., Hunt, D.F., et al. (2000). Mitotic phosphorylation of histone H3 is governed by Ipl1/aurora kinase and Glc7/PP1 phosphatase in budding yeast and nematodes. Cell 102, 279–291. Article
Hyman, A.A., and White, J.G. (1987). Determination of cell division axes in the early embryogenesis of Caenorhabditis elegans. J. Cell Biol. 105, 2123–2135. Abstract Article
Jantsch-Plunger, V., and Glotzer, M. (1999). Depletion of syntaxins in the early Caenorhabditis elegans embryo reveals a role for membrane fusion events in cytokinesis. Curr. Biol. 9, 738–745. Abstract Article
Jantsch-Plunger, V., Gönczy, P., Romano, A., Schnabel, H., Hamill, D., Schnabel, R., Hyman, A.A., and Glotzer, M. (2000). CYK-4: A Rho family GTPase activating protein (GAP) required for central spindle formation and cytokinesis. J. Cell Biol. 149, 1391–1404. Abstract Article
Kaitna, S., Mendoza, M., Jantsch-Plunger, V., and Glotzer, M. (2000). Incenp and an aurora-like kinase form a complex essential for chromosome segregation and efficient completion of cytokinesis. Curr. Biol. 10, 1172–1181. Abstract Article
Kaitna, S., Pasierbek, P., Jantsch, M., Loidl, J., and Glotzer, M. (2002). The aurora B kinase AIR-2 regulates kinetochores during mitosis and is required for separation of homologous Chromosomes during meiosis. Curr. Biol. 12, 798–812. Abstract Article
Kamath, R.S., Fraser, A.G., Dong, Y., Poulin, G., Durbin, R., Gotta, M., Kanapin, A., Le Bot, N., Moreno, S., Sohrmann, M., et al. (2003). Systematic functional analysis of the Caenorhabditis elegans genome using RNAi. Nature 421, 231–237. Abstract Article
Kemp, C.A., Kopish, K.R., Zipperlen, P., Ahringer, J., and O'Connell, K.F. (2004). Centrosome maturation and duplication in C. elegans require the coiled-coil protein SPD-2. Dev. Cell 6, 511–523. Abstract Article
Kemphues, K.J., Kusch, M., and Wolf, N. (1988). Maternal-effect lethal mutations on linkage group II of Caenorhabditis elegans. Genetics 120, 977–986. Abstract
Khodjakov, A., Cole, R.W., Oakley, B.R., and Rieder, C.L. (2000). Centrosome-independent mitotic spindle formation in vertebrates. Curr. Biol. 10, 59–67. Abstract Article
Kimura, A., and Onami, S. (2005). Computer Simulations and Image Processing Reveal Length-Dependent Pulling Force as the Primary Mechanism for C. elegans Male Pronuclear Migration. Developmental Cell 8, 765–775. Article
Kirkham, M., Muller-Reichert, T., Oegema, K., Grill, S., and Hyman, A.A. (2003). SAS-4 is a C. elegans centriolar protein that controls centrosome size. Cell 112, 575–587. Abstract Article
Kitagawa, R., and Rose, A.M. (1999). Components of the spindle-assembly checkpoint are essential in Caenorhabditis elegans. Nat. Cell Biol. 1, 514–521. Article
Kuznetsov, N.V., Sandblad, L., Hase, M.E., Hunziker, A., Hergt, M., and Cordes, V.C. (2002). The evolutionarily conserved single-copy gene for murine Tpr encodes one prevalent isoform in somatic cells and lacks paralogs in higher eukaryotes. Chromosoma 111, 236–255. Abstract Article
Labbe, J.C., McCarthy, E.K., and Goldstein, B. (2004). The forces that position a mitotic spindle asymmetrically are tethered until after the time of spindle assembly. J. Cell Biol. 167, 245–256. Abstract Article
Le Bot, N., Tsai, M.C., Andrews, R.K., and Ahringer, J. (2003). TAC-1, a regulator of microtubule length in the C. elegans embryo. Curr. Biol. 13, 1499–1505. Abstract Article
Lee, K.K., Gruenbaum, Y., Spann, P., Liu, J., and Wilson, K.L. (2000). C. elegans nuclear envelope proteins emerin, MAN1, lamin, and nucleoporins reveal unique timing of nuclear envelope breakdown during mitosis. Mol. Biol. Cell 11, 3089–3099. Abstract
Lee, K.K., Starr, D., Cohen, M., Liu, J., Han, M., Wilson, K.L., and Gruenbaum, Y. (2002). Lamin-dependent localization of UNC-84, a protein required for nuclear migration in Caenorhabditis elegans. Mol. Biol. Cell 13, 892–901. Abstract Article
Lee, K.K., and Wilson, K.L. (2004). All in the family: evidence for four new LEM-domain proteins Lem2 (NET-25), Lem3, Lem4 and Lem5 in the human genome. Symp. Soc. Exp. Biol. 329–339. Abstract
Leidel, S., Delattre, M., Cerutti, L., Baumer, K., and Gönczy, P. (2005). SAS-6 defines a protein family required for centrosome duplication in C. elegans and in human cells. Nat. Cell Biol. 7, 115–125. Abstract Article
Leidel, S., and Gönczy, P. (2003). SAS-4 is essential for centrosome duplication in C elegans and is recruited to daughter centrioles once per cell cycle. Dev. Cell 4, 431–439. Abstract Article
Lieb, J.D., Albrecht, M.R., Chuang, P.T., and Meyer, B.J. (1998). MIX-1: an essential component of the C. elegans mitotic machinery executes X chromosome dosage compensation. Cell 92, 265–277. Article
Lin, F., Blake, D.L., Callebaut, I., Skerjanc, I.S., Holmer, L., McBurney, M.W., Paulin-Levasseur, M., and Worman, H.J. (2000). MAN1, an inner nuclear membrane protein that shares the LEM domain with lamina-associated polypeptide 2 and emerin. J. Biol. Chem. 275, 4840–4847. Abstract Article
Liu, J., Ben-Shahar, T.R., Riemer, D., Treinin, M., Spann, P., Weber, K., Fire, A., and Gruenbaum, Y. (2000). Essential roles for Caenorhabditis elegans lamin gene in nuclear organization, cell cycle progression, and spatial organization of nuclear pore complexes. Mol. Biol. Cell 11, 3937–3947. Abstract
Liu, J., Lee, K.K., Segura-Totten, M., Neufeld, E., Wilson, K.L., and Gruenbaum, Y. (2003). MAN1 and emerin have overlapping function(s) essential for chromosome segregation and cell division in Caenorhabditis elegans. Proc. Natl. Acad. Sci. USA 100, 4598–4603. Abstract Article
Maddox, A.S., Habermann, B., Desai, A., and Oegema, K. (2005). Distinct roles for two C. elegans anillins in the gonad and early embryo. Development 132, 2837–2848. Abstract Article
Maddox, P.S., Oegema, K., Desai, A., and Cheeseman, I.M. (2004). "Holo"er than thou: chromosome segregation and kinetochore function in C. elegans. Chromosome Res. 12, 641–653. Abstract Article
Maeda, I., Kohara, Y., Yamamoto, M., and Sugimoto, A. (2001). Large-scale analysis of gene function in Caenorhabditis elegans by high-throughput RNAi. Curr. Biol. 11, 171–176. Abstract Article
Malone, C.J., Fixsen, W.D., Horvitz, H.R., and Han, M. (1999). UNC-84 localizes to the nuclear envelope and is required for nuclear migration and anchoring during C. elegans development. Development 126, 3171–3181.
Malone, C.J., Misner, L., Le Bot, N., Tsai, M.C., Campbell, J.M., Ahringer, J., and White, J.G. (2003). The C. elegans hook protein, ZYG-12, mediates the essential attachment between the centrosome and nucleus. Cell 115, 825–836. Abstract Article
Margalit, A., Segura-Totten, M., Gruenbaum, Y., and Wilson, K.L. (2005). Barrier-to-autointegration factor is required to segregate and enclose chromosomes within the nuclear envelope and assemble the nuclear lamina. Proc. Natl. Acad. Sci. USA 102, 3290–3295. Abstract Article
Matthews, L.R., Carter, P., Thierry-Mieg, D., and Kemphues, K. (1998). ZYG-9, a Caenorhabditis elegans protein required for microtubule organization and function, is a component of meiotic and mitotic spindle poles. J. Cell Biol. 141, 1159–1168. Abstract Article
Megraw, T.L., Li, K., Kao, L.R., and Kaufman, T.C. (1999). The centrosomin protein is required for centrosome assembly and function during cleavage in Drosophila. Development 126, 2829–2839. Abstract
Mishima, M., Kaitna, S., and Glotzer, M. (2002). Central spindle assembly and cytokinesis require a kinesin-like protein/RhoGAP complex with microtubule bundling activity. Dev. Cell 2, 41–54. Abstract Article
Mito, Y., Sugimoto, A., and Yamamoto, M. (2003). Distinct developmental function of two Caenorhabditis elegans homologs of the cohesin subunit Scc1/Rad21. Mol. Biol. Cell 14, 2399–2409. Abstract Article
Moore, L.L., Morrison, M., and Roth, M.B. (1999). HCP-1, a protein involved in chromosome segregation, is localized to the centromere of mitotic chromosomes in Caenorhabditis elegans. J. Cell Biol. 147, 471–480. Abstract Article
Moore, L.L., and Roth, M.B. (2001). HCP-4, a CENP-C-like protein in Caenorhabditis elegans, is required for resolution of sister centromeres. J. Cell Biol. 153, 1199–1208. Abstract Article
Moore, L.L., Stanvitch, G., Roth, M.B., and Rosen, D. (2005). HCP-4/CENP-C promotes the prophase timing of centromere resolution by enabling the centromere association of HCP-6 in Caenorhabditis elegans. Mol. Cell Biol. 25, 2583–2592. Abstract Article
Munro, E., Nance, J., and Priess, J.R. (2004). Cortical flows powered by asymmetrical contraction transport PAR proteins to establish and maintain anterior-posterior polarity in the early C. elegans embryo. Dev. Cell 7, 413–424. Abstract Article
Nguyen, T.Q., Sawa, H., Okano, H., and White, J.G. (2000). The C. elegans septin genes, unc-59 and unc-61, are required for normal postembryonic cytokineses and morphogenesis but have no essential function in embryogenesis. J. Cell Sci. 113(Pt 21), 3825–3837.
Nystul, T.G., Goldmark, J.P., Padilla, P.A., and Roth, M.B. (2003). Suspended animation in C. elegans requires the spindle checkpoint. Science 302, 1038–1041. Article
Nystul, T.G., and Roth, M.B. (2004). Carbon monoxide-induced suspended animation protects against hypoxic damage in Caenorhabditis elegans. Proc. Natl. Acad. Sci. USA 101, 9133–9136. Article
O'Connell, K.F. (2000). The centrosome of the early C. elegans embryo: inheritance, assembly, replication, and developmental roles. Curr. Top. Dev. Biol. 49, 365–384. Abstract
O'Connell, K.F., Caron, C., Kopish, K.R., Hurd, D.D., Kemphues, K.J., Li, Y., and White, J.G. (2001). The C. elegans zyg-1 gene encodes a regulator of centrosome duplication with distinct maternal and paternal roles in the embryo. Cell 105, 547–558. Article
O'Connell, K.F., Leys, C.M., and White, J.G. (1998). A genetic screen for temperature-sensitive cell-division mutants of Caenorhabditis elegans. Genetics 149, 1303–1321. Abstract
O'Connell, K.F., Maxwell, K.N., and White, J.G. (2000). The spd-2 gene is required for polarization of the anteroposterior axis and formation of the sperm asters in the Caenorhabditis elegans zygote. Dev. Biol. 222, 55–70. Abstract Article
O'Toole, E.T., McDonald, K.L., Mantler, J., McIntosh, J.R., Hyman, A.A., and Müller-Reichert, T. (2003). Morphologically distinct microtubule ends in the mitotic centrosome of Caenorhabditis elegans. J. Cell Biol. 163, 451–456. Abstract Article
Oegema, K., Desai, A., Rybina, S., Kirkham, M., and Hyman, A.A. (2001). Functional analysis of kinetochore assembly in Caenorhabditis elegans. J. Cell Biol. 153, 1209–1226. Abstract Article
Ono, K., Parast, M., Alberico, C., Benian, G.M., and Ono, S. (2003). Specific requirement for two ADF/cofilin isoforms in distinct actin-dependent processes in Caenorhabditis elegans. J. Cell Sci. 116, 2073–2085. Abstract Article
Ono, T., Losada, A., Hirano, M., Myers, M.P., Neuwald, A.F., and Hirano, T. (2003). Differential contributions of condensin I and condensin II to mitotic chromosome architecture in vertebrate cells. Cell 115, 109–121. Article
Palazzo, R.E., Vogel, J.M., Schnackenberg, B.J., Hull, D.R., and Wu, X. (2000). Centrosome maturation. Curr. Top. Dev. Biol. 49, 449–470. Abstract
Pasierbek, P., Fodermayr, M., Jantsch, V., Jantsch, M., Schweizer, D., and Loidl, J. (2003). The Caenorhabditis elegans SCC-3 homologue is required for meiotic synapsis and for proper chromosome disjunction in mitosis and meiosis. Exp. Cell Res. 289, 245–255. Abstract Article
Pelletier, L., Müller-Reicher, T., Srayko, M., Ozlu, N., Schlaitz, A., and Hyman, A.A. (2004). The C. elegans centrosome during early embryonic development. In Centrosomes in Development and Disease, E. Nigg, ed. (Weinheim, Wiley-VCH). Abstract Article
Pelletier, L., Ozlu, N., Hannak, E., Cowan, C., Habermann, B., Ruer, M., Müller-Reichert, T., and Hyman, A.A. (2004). The Caenorhabditis elegans centrosomal protein SPD-2 is required for both pericentriolar material recruitment and centriole duplication. Curr. Biol. 14, 863–873. Article
Piano, F., Schetter, A.J., Mangone, M., Stein, L., and Kemphues, K.J. (2000). RNAi analysis of genes expressed in the ovary of Caenorhabditis elegans. Curr. Biol. 10, 1619–1622. Abstract Article
Piekny, A.J., and Mains, P.E. (2002). Rho-binding kinase (LET-502) and myosin phosphatase (MEL-11) regulate cytokinesis in the early Caenorhabditis elegans embryo. J. Cell Sci. 115, 2271–2282. Abstract
Powers, J., Bossinger, O., Rose, D., Strome, S., and Saxton, W. (1998). A nematode kinesin required for cleavage furrow advancement. Curr. Biol. 8, 1133–1136. Abstract Article
Powers, J., Rose, D.J., Saunders, A., Dunkelbarger, S., Strome, S., and Saxton, W.M. (2004). Loss of KLP-19 polar ejection force causes misorientation and missegregation of holocentric chromosomes. J. Cell Biol. 166, 991–1001. Abstract Article
Praitis, V., Casey, E., Collar, D., and Austin, J. (2001). Creation of low-copy integrated transgenic lines in Caenorhabditis elegans. Genetics 157, 1217–1226. Abstract
Preble, A.M., Giddings, T.M., Jr., and Dutcher, S.K. (2000). Basal bodies and centrioles: their function and structure. Curr. Top Dev. Biol. 49, 207–233. Abstract
Raich, W.B., Moran, A.N., Rothman, J.H., and Hardin, J. (1998). Cytokinesis and midzone microtubule organization in Caenorhabditis elegans require the kinesin-like protein ZEN-4. Mol. Biol. Cell 9, 2037–2049. Abstract
Rebollo, E., Llamazares, S., Reina, J., and Gonzalez, C. (2004). Contribution of noncentrosomal microtubules to spindle assembly in Drosophila spermatocytes. PLoS Biol. 2, E8. Abstract Article
Riemer, D., Dodemont, H., and Weber, K. (1993). A nuclear lamin of the nematode Caenorhabditis elegans with unusual structural features; cDNA cloning and gene organization. Eur. J. Cell Biol. 62, 214–223. Abstract
Rogers, E., Bishop, J.D., Waddle, J.A., Schumacher, J.M., and Lin, R. (2002). The aurora kinase AIR-2 functions in the release of chromosome cohesion in Caenorhabditis elegans meiosis. J. Cell Biol. 157, 219–229. Abstract Article
Romano, A., Guse, A., Krascenicova, I., Schnabel, H., Schnabel, R., and Glotzer, M. (2003). CSC-1: a subunit of the Aurora B kinase complex that binds to the survivin-like protein BIR-1 and the incenp-like protein ICP-1. J. Cell Biol. 161, 229–236. Abstract Article
Rual, J.F., Ceron, J., Koreth, J., Hao, T., Nicot, A.S., Hirozane-Kishikawa, T., Vandenhaute, J., Orkin, S.H., Hill, D.E., van den Heuvel, S., and Vidal, M. (2004). Toward improving Caenorhabditis elegans phenome mapping with an ORFeome-based RNAi library. Genome Res. 14, 2162–2168. Abstract Article
Scaerou, F., Starr, D.A., Piano, F., Papoulas, O., Karess, R.E., and Goldberg, M.L. (2001). The ZW10 and Rough Deal checkpoint proteins function together in a large, evolutionarily conserved complex targeted to the kinetochore. J. Cell Sci. 114, 3103–3114.
Schmidt, D.J., Rose, D.J., Saxton, W.M., and Strome, S. (2005). Functional analysis of cytoplasmic dynein heavy chain in Caenorhabditis elegans with fast-acting temperature-sensitive mutations. Mol. Biol. Cell 16, 1200–1212. Abstract Article
Schmutz, C., and Spang, A. (2005). Knockdown of the centrosomal component SAS-5 results in defects in nuclear morphology in Caenorhabditis elegans. Eur. J. Cell Biol. 84, 75–82. Abstract Article
Schneider, S.Q., and Bowerman, B. (2003). Cell polarity and the cytoskeleton in the Caenorhabditis elegans zygote. Annu. Rev. Genet. 37, 221–249. Abstract Article
Schumacher, J.M., Ashcroft, N., Donovan, P.J., and Golden, A. (1998). A highly conserved centrosomal kinase, AIR-1, is required for accurate cell cycle progression and segregation of developmental factors in Caenorhabditis elegans embryos. Development 125, 4391–4402. Abstract
Schumacher, J.M., Golden, A., and Donovan, P.J. (1998). AIR-2: An Aurora/Ipl1-related protein kinase associated with chromosomes and midbody microtubules is required for polar body extrusion and cytokinesis in Caenorhabditis elegans embryos. J. Cell Biol. 143, 1635–1646. Abstract Article
Segura-Totten, M., and Wilson, K.L. (2004). BAF: roles in chromatin, nuclear structure and retrovirus integration. Trends Cell Biol. 14, 261–266. Abstract Article
Severson, A.F., Baillie, D.L., and Bowerman, B. (2002). A Formin Homology protein and a profilin are required for cytokinesis and Arp2/3-independent assembly of cortical microfilaments in C. elegans. Curr. Biol. 12, 2066–2075. Abstract Article
Severson, A.F., Hamill, D.R., Carter, J.C., Schumacher, J., and Bowerman, B. (2000). The aurora-related kinase AIR-2 recruits ZEN-4/CeMKLP1 to the mitotic spindle at metaphase and is required for cytokinesis. Curr. Biol. 10, 1162–1171. Abstract Article
Shelton, C.A., Carter, J.C., Ellis, G.C., and Bowerman, B. (1999). The nonmuscle myosin regulatory light chain gene mlc-4 is required for cytokinesis, anterior-posterior polarity, and body morphology during Caenorhabditis elegans embryogenesis. J. Cell Biol. 146, 439–451. Abstract Article
Simmer, F., Moorman, C., van der Linden, A.M., Kuijk, E., van den Berghe, P.V., Kamath, R.S., Fraser, A.G., Ahringer, J., and Plasterk, R.H. (2003). Genome-wide RNAi of C. elegans using the hypersensitive rrf-3 strain reveals novel gene functions. PLoS Biol. 1, E12. Abstract Article
Skop, A.R., Bergmann, D., Mohler, W.A., and White, J.G. (2001). Completion of cytokinesis in C. elegans requires a brefeldin A-sensitive membrane accumulation at the cleavage furrow apex. Curr. Biol. 11, 735–746. Abstract Article
Sõnnichsen, B., Koski, L.B., Walsh, A., Marschall, P., Neumann, B., Brehm, M., Alleaume, A.M., Artelt, J., Bettencourt, P., Cassin, E., et al. (2005). Full-genome RNAi profiling of early embryogenesis in Caenorhabditis elegans. Nature 434, 462–469. Abstract Article
Speliotes, E.K., Uren, A., Vaux, D., and Horvitz, H.R. (2000). The survivin-like C. elegans BIR-1 protein acts with the Aurora-like kinase AIR-2 to affect chromosomes and the spindle midzone. Mol. Cell 6, 211–223. Abstract Article
Srayko, M., Quintin, S., Schwager, A., and Hyman, A.A. (2003). Caenorhabditis elegans TAC-1 and ZYG-9 form a complex that is essential for long astral and spindle microtubules. Curr. Biol. 13, 1506–1511. Abstract Article
Starr, D.A., and Han, M. (2002). Role of ANC-1 in tethering nuclei to the actin cytoskeleton. Science 298, 406–409. Article
Starr, D.A., Hermann, G.J., Malone, C.J., Fixsen, W., Priess, J.R., Horvitz, H.R., and Han, M. (2001). unc-83 encodes a novel component of the nuclear envelope and is essential for proper nuclear migration. Development 128, 5039–5050.
Starr, D.A., Williams, B.C., Hays, T.S., and Goldberg, M.L. (1998). ZW10 helps recruit dynactin and dynein to the kinetochore. J. Cell Biol. 142, 763–774. Article
Stear, J.H., and Roth, M.B. (2002). Characterization of HCP-6, a C. elegans protein required to prevent chromosome twisting and merotelic attachment. Genes Dev. 16, 1498–1508. Article
Stear, J.H., and Roth, M.B. (2004). The Caenorhabditis elegans kinetochore reorganizes at prometaphase and in response to checkpoint stimuli. Mol. Biol. Cell 15, 5187–5196. Article
Strome, S., Powers, J., Dunn, M., Reese, K., Malone, C.J., White, J., Seydoux, G., and Saxton, W. (2001). Spindle dynamics and the role of γ-tubulin in early Caenorhabditis elegans embryos. Mol. Biol. Cell 12, 1751–1764. Abstract
Strome, S., and Wood, W.B. (1983). Generation of asymmetry and segregation of germ-line granules in early C. elegans embryos. Cell 35, 15–25. Abstract Article
Sullivan, K.F. (2001). A solid foundation: functional specialization of centromeric chromatin. Curr. Opin. Genet. Dev. 11, 182–188. Abstract Article
Swan, K.A., Severson, A.F., Carter, J.C., Martin, P.R., Schnabel, H., Schnabel, R., and Bowerman, B. (1998). cyk-1: a C. elegans FH gene required for a late step in embryonic cytokinesis. J. Cell Sci. 111(Pt 14), 2017–2027. Abstract
Thompson, H.M., Skop, A.R., Euteneuer, U., Meyer, B.J., and McNiven, M.A. (2002). The large GTPase dynamin associates with the spindle midzone and is required for cytokinesis. Curr. Biol. 12, 2111–2117. Abstract Article
Tsou, M.F., Hayashi, A., DeBella, L.R., McGrath, G., and Rose, L.S. (2002). LET-99 determines spindle position and is asymmetrically enriched in response to PAR polarity cues in C. elegans embryos. Development 129, 4469–4481. Abstract
Vagnarelli, P., and Earnshaw, W.C. (2004). Chromosomal passengers: the four-dimensional regulation of mitotic events. Chromosoma 113, 211–222. Abstract Article
Vaizel-Ohayon, D., and Schejter, E.D. (1999). Mutations in centrosomin reveal requirements for centrosomal function during early Drosophila embryogenesis. Curr. Biol. 9, 889–898. Abstract Article
Verbrugghe, K.J., and White, J.G. (2004). SPD-1 is required for the formation of the spindle midzone but is not essential for the completion of cytokinesis in C. elegans embryos. Curr. Biol. 14, 1755–1760. Abstract Article
Walther, T.C., Askjaer, P., Gentzel, M., Habermann, A., Griffiths, G., Wilm, M., Mattaj, I.W., and Hetzer, M. (2003). RanGTP mediates nuclear pore complex assembly. Nature 424, 689–694. Abstract Article
Wolf, N., Hirsh, D., and McIntosh, J.R. (1978). Spermatogenesis in males of the free-living nematode, Caenorhabditis elegans. J. Ultrastruct. Res. 63, 155–169. Abstract Article
Yang, H.Y., McNally, K., and McNally, F.J. (2003). MEI-1/katanin is required for translocation of the meiosis I spindle to the oocyte cortex in C. elegans. Dev. Biol. 260, 245–259. Abstract Article
*Edited by James M. Kramer and Donald G. Moerman. Last revised May 4, 2005. Published January 19, 2006. This chapter should be cited as: Oegema, K. and Hyman, A. A. Cell division (January 19, 2006), WormBook, ed. The C. elegans Research Community, WormBook, doi/10.1895/wormbook.1.72.1, http://www.wormbook.org.
Copyright: © 2006 Karen Oegema and Anthony A. Hyman. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
§To whom correspondence should be addressed. E-mail: [email protected] or [email protected]
 All WormBook content, except where otherwise noted, is licensed under a Creative Commons Attribution License.
All WormBook content, except where otherwise noted, is licensed under a Creative Commons Attribution License.